Развитие и изменения эндоплазматического ретикулума (ЭПР)
• Развитие ЭПР и состав его мембран меняются в зависимости от запросов клетки
• Перемещения мембран ЭПР происходят вдоль цитоскелета
• Механизм расширения, сокращения и образования трубочек ЭПР пока не выяснен
• Сигнальные процессы, контролирующие состав ЭПР до конца не выяснены, однако они могут перекрываться с откликом неструктурированных белков
Подобно другим органеллам, эндоплазматический ретикулум (ЭПР) должен адаптироваться к нуждам клетки. При необходимости отдельные регионы ЭПР должны расширяться или сокращаться. Клетка должна иметь возможность собирать и реорганизовывать органеллу, а также поддерживать ее форму. При митозе ЭПР должен распределяться между дочерними клетками. Хотя в отношении молекулярных основ этих процессов известно немного, очевидно, что органелла характеризуется высокой динамичностью.
Форма эндоплазматического ретикулума (ЭПР) постоянно меняется. Особенно это касается трубочек эндоплазматического ретикулума (ЭПР), которые находятся в постоянном движении: они удлиняются, сокращаются, разветвляются и сливаются между собой. Это движение определенным образом связано с цитоскелетом; в клетках млекопитающих трубочки ЭПР часто ориентированы в соответствии с микротрубочками, так, что их расширение и сокращение происходят вдоль этих структур цитоскелета. В клетках дрожжей ЭПР часто связан с актиновыми филаментами.
Однако разрушение цитоскелета не приводит к дисфункции эндоплазматического ретикулума (ЭПР) или к коллапсу системы его трубочек. Для образования трубочек или их сетей in vitro не требуются элементы цитоскелета. Поэтому представляется вероятным, что образование трубочек и их сборка в сеть являются характерным свойством ЭПР, которое не зависит от цитоскелета. Скорее, роль цитоскелета может заключаться в том, чтобы обеспечить надлежащее распределение сети трубочек в клетке после ее сборки.
Механизмы, посредством которых происходит сборка новых или расширение существующих мембран эндоплазматического ретикулума (ЭПР), неизвестны, однако выяснены некоторые процессы, играющие ключевую роль. Образование трубочек ЭПР in vitro требует АТФ и ГТФ, а также, возможно, белка, близкого к участвующему в слиянии везикул ЭПР-Гольджи (белок NSF). Это позволяет предполагать, что пролиферация ЭПР включает события слияния, при котором из маленьких везикул ЭПР образуются большие разветвленные цепи взаимосвязанных везикул.
Для того чтобы при слиянии образовались трубочки, а не аморфные массы, процесс должен определенным образом регулироваться. У всех эукариот преобладает тубулярная форма ЭПР, что позволяет предполагать, что формообразование является контролируемым и функционально важным процессом. Тубулярная форма может отчасти возникать за счет селективного распределения липидов в местах их локализации при расширении сети ЭПР. У молекул некоторых липидов головки более крупные, чем хвостовые участки, и в результате их присутствие искривляет бислой. Расположение этих липидов может вызывать локальное скручивание мембраны, благоприятствующее образованию трубочек.
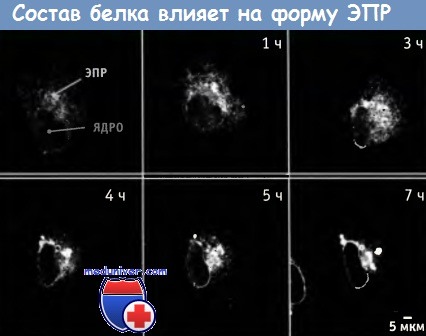
Гиперэкспрессия флуоресцирующего трансмембранного белка на протяжении нескольких часов приводит к изменению структуры эндоплазматического ретикулума (ЭПР).
Еще одна возможность состоит в том, что трубочки ЭПР формируются за счет белкового каркаса, находящегося на одной из поверхностей мембраны, который и обеспечивает необходимую кривизну. По крайней мере, частично, форма ЭПР зависит от содержания в нем белков. На рисунке ниже показано, как слабые взаимодействия между трансмембранными белками, входящими в состав ЭПР, значительно меняют форму и характер распределения этой органеллы в клетке. Остается неясным, каким образом плоские слои, характерные для некоторых гранулярных ЭПР, принимают свою форму и почему они часто группируются в структуры, напоминающие стопки.
Необходимость регуляции размера и формы ЭПР подтверждается поведением этой органеллы в клетках, которые предъявляют повышенные требования к сети ЭПР. У клеток, выполняющих секреторные функции, например эндокринных, развивается протяженный гранулярный ЭПР. Это достигается за счет механизма обратной связи, который сигнализирует о необходимости усиления синтеза липидов и белков ЭПР в ответ на усиление транспорта белков через ЭПР. Препараты, которые обезвреживаются в гладком ЭПР, например фенобарбитал, также индуцируют развитие органеллы, и их выведение быстро возвращает гладкий ЭПР к исходному состоянию.
Еще одно загадочное проявление способности ЭПР изменять размер, состоит в образовании кармеллы при гиперэкспрессии некоторых мембранных белков. На рисунке ниже приведена клетка, в которой образовалась кармелла. Эта структура представляет собой образование из стопок мембран ЭПР, тесно связанное с ядерной оболочкой и заполненное белком. Вероятно, кармеллы служат резервуарами, в которых хранится избыток белков. Опять-таки, возможно существование механизма обратной связи между ЭПР и аппаратами генной экспрессии и синтеза липидов.
Интересно, что отклик неструктурированных белков, представляющий собой одну из систем сигнализации между ЭПР и ядром, о которой говорилось выше, может играть роль в пролиферации ЭПР. Активация UPR не только стимулирует синтез шаперонов в ЭПР, но также синтез инозитола. Последний представляет собой головку молекулы фосфатидилинозитола, одного из четырех основных липидов мембран. У дрожжей эта активация происходит с участием белков Ire1p и Hac1p, т. е. таким же образом, как и активация синтеза шаперонов. Для обеспечения развития новых мембран ЭПР также активируется синтез других фосфолипидов. Таким образом, UPR может обусловливать запуск каскада процессов, участвующих в перестройках ЭПР. Хотя это предположение достаточно привлекательно, оно еще не доказано.
Какой механизм дает клетке возможность воспринимать поступающий при стрессе сигнал как свидетельствующий о необходимости пролиферации ЭПР, а не как сигнал к гибели? При дифференцировке В-клеток в плазматические клетки, секретирующие антитела, активация UPR и развитие мембран ЭПР происходят до усиления синтеза иммуноглобулинов. Эти данные позволяют предполагать, что существует механизмы, по которым UPR активируется раньше, чем возрастает нагрузка на ЭПР, а не в ответ на нее. Аналогичным образом, ЭПР-компартменты у растений интенсивно используются для хранения различных белков и масел, т. е. компонентов, важных для развития семян и для защиты от патогенов. Эти белки могут агрегировать в компартментах ЭПР, но таким способом, который не вызывает их деградации. Накоплению белков в таких компартментах также предшествует активация синтеза шаперонов ЭПР.
Еще один процесс, иллюстрирующий реструктуризацию органеллы, представляет собой распределение ЭПР при митозе. При этом данная органелла не синтезируется de novo, а происходит из предсуществующего ЭПР, поэтому важно обеспечить распределение ее по дочерним клеткам. Одинаковое распределение между дочерними клетками имеет свои преимущества. Когда одна клетка получает значительно меньше половины массы ЭПР, то происходит задержка в следующем клеточном цикле, которая во многих ситуациях представляет опасность. В наибольшей степени подготовка ЭПР к митозу проявляется в области ядерной оболочки. С началом митоза она фрагментируется, и происходит диссоциация комплексов ядерных пор.
Эти процессы позволяют предполагать, что существуют сигналы разрушения ядерной оболочки, предшествующие митозу, и сигналы последующей ее сборки. Остальная часть ЭПР также должна сегрегировать при митозе. В некоторых клетках эта сегрегация сопровождается везикуляризацией (т. е. распадом) и повторным образованием ЭПР после завершения митоза. Однако в большинстве клеток в течение митоза ЭПР поддерживается в виде целостной сети и делится на две примерно равные части при цитокинезе. В некоторых случаях, например в раннем эмбриогенезе, ассоциация ЭПР с микротрубочками митотического веретена помогает равномерному распределению ЭПР между дочерними клетками.
Еще один аспект биогенеза ЭПР, который в значительной степени остается неисследованным, представляет собой слияние друг с другом компонентов ЭПР. (Это гомотопическое слияние, поскольку две мембраны принад лежат одной и той же органелле). Гомотопическое слияние необходимодля образования сети ЭПР, для сборки ядерной оболочки после митоза, и происходит при слиянии ядер двух гаплоидных клеток, что у дрожжей известно как кариогамия Для протекания кариогамии необходимы, видимо, те же белки, что и для слияния ЭПР-Голвджи (NSFs, SNAREs, АТФазы и несколько G-белков). Кариогамия уникальна в том отношении, что для нее требуется несколько компонентов поспрансля-ционного транслокона, однако отличных от тех, которые образуют канал.
(Белок KAR2, представляющий собой дрожжевой гомолог BiP, был сначала идентифицирован как участник кариогамии). Какую роль играют в кариогамии эти белки, пока неизвестно. Согласно одному из представлений, они непосредственно не связаны с кариогамией, а способствуют транслокации других белков-участников этого процесса.
Гиперэкспрессия некоторых трансмембранных белков приводит к образованию стопок ЭПР, называемых кармеллами.
Представлена микрофотография дрожжевой клетки в электронном микроскопе, на которой видны кармеллы, окружающие ядро.
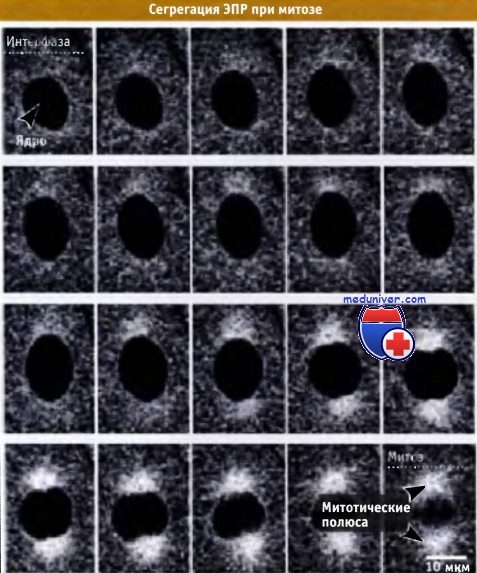
Флуоресцентная микрофотография ЭПР бластомера морского ежа при вхождении в митоз.
Интерфаза представлена на левом верхнем снимке, а метафаза — внизу справа. При делении ЭПР сохраняется в виде непрерывной сети и концентрируется на полюсах.
Экспрессия флуоресцирующего белка, локализованного в ЭПР, дает возможность наблюдать за этим процессом.