Механизм возвращения в цитозоль и деградации неправильно свернутых белков клетки - ERAD
• Транслоцированные в ЭПР белки могут экспортироваться обратно в цитозоль, где они подвергаются убиквитинилированию и деградации протеосомами — процесс известный как деградация, связанная с ЭПР.
• Белки возвращаются в цитозоль путем ретроградной транслокации, механизм которой менее понятен, чем механизм транслокации в ЭПР.
Контроль за структурой белков, осуществляемый в ЭПР, обеспечивает экспорт только таких трансмембранных и секреторных белков, которые обладают правильной нативной конфигурацией, в то время как белки неправильной структуры удерживаются в ЭПР шаперонами и имеют шанс исправить конфигурацию. Однако что же происходит с белками, которым не удается приобрести правильную структуру?
Такие белки должны каким-то образом деградировать. Одним из путей решения этой проблемы служит использование ретроградной транслокации (иногда называемой дислокация или ретротранслокация) для экспорта неправильно собранных белков обратно в цитозоль. Там они взаимодействуют с убиквитином и деградируют с участием больших протеазных комплексов, протеосом. Этот путь деградации называется — деградация, связанная с ЭПР (ERAD).
Открытие этого пути последовало после понимания того факта, что белки, обладающие неправильной нативной структурой, не накапливаются в ЭПР, а деградируют. Первоначально предполагалось, что деградация происходит в самом ЭПР, однако, несмотря на тщательные поиски, в этой органелле не были обнаружены соответствующие протеазы. Вместо этого начали появляться данные о том, что белки деградируют под действием протеаз, содержащихся в цитозоле.
Вначале было показано, что химические ингибиторы протеосом блокируют деградацию вновь синтезированного интегрального мембранного белка (CFTR, или регулятор трансмембранного переноса при кистозном фиброзе). Эти данные позволили предположить, что белки должны экспортироваться из ЭПР до деградации. В дальнейшем было показано, что ингибирование функций протеосом приводит к накоплению в цитозоле полностью транслоцированных белков.
Каким образом белок отбирается для ERAD? В отличие от процесса транслокации в ЭПР сигнальная последовательность на белке для его направления на деградацию отсутствует. Скорее всего, при ERAD распознаются какие-то свойства, которые отличают белки с неправильной структурой от белков, обладающих нативной конформацией или временно не имеющих правильной структуры высшего порядка.
Например, мутация, которая дестабилизирует нативную структуру белка, может вызывать ее деградацию. Фактически мутировавший белок может быть признан субстратом для ERAD, даже если мутация мало повлияла на его функцию. Даже белки, не обладающие заметными дефектами структуры, могут быть отобраны для ERAD, что свидетельствует о существовании скрытых, неизвестных нам признаков, которые, однако, узнаются клеткой.
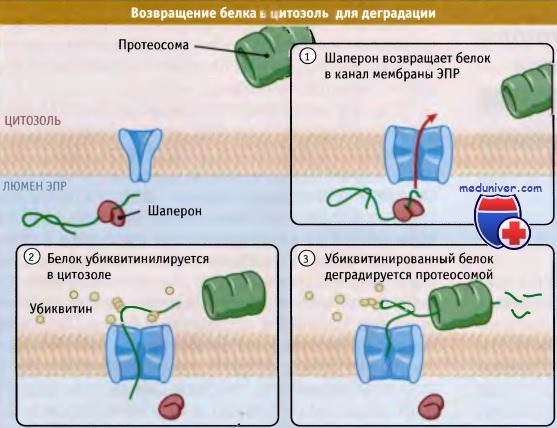
Рисунок ниже иллюстрирует основные этапы ERAD. Первый этап отбора белка для ERAD — это обнаружение неправильной структуры. Хотя мы и не знаем, каким образом это происходит, возможно, что существенную роль здесь играют шапероны. Однако ассоциация с шаперонами не может являться единственным критерием отбора белка для ERAD, поскольку шапероны участвуют в организации конформации всех новообразующихся белков. Не исключено, что сигнал к ретроградной транслокации зависит от промежутка времени, в течение которого белок находится в связанной с шаперонами форме.
При ретроградной транслокации белки с неправильной нативной структурой возвращаются из люмена эндоплазматического ретикулума (ЭПР) обратно в его мембрану и в цитоплазму.
Там они подвергаются деградации в протеосомах. По-видимому, определенную роль в переадресовании играют шапероны люмена,
поскольку они должны связываться с белками, обладающими неправильной нативной структурой.
Шапероны остаются в связанной форме до тех пор, пока белок не приобретет нативную структуру. Если образование такой структуры невозможно, то белок дольше, чем обычно, будет оставаться связанным с шаперонами. Впрочем, вообще не ясно, каким образом продолжительность связи белка с шаперонами дает сигнал к ретроградной транслокации. В случае неправильной структуры гликопротеинов фермент а-маннозидаза I отщепляет маннозу и тем самым дает сигнал к деградации белка. Не ясно, однако, что служит сигналом к отщеплению маннозы.
После отбора белка для ретроградной транслокации, он должен пройти через мембрану ЭПР. Многочисленные генетические и биохимические данные позволяют предполагать, что роль канала ретроградной транслокации выполняет комплекс Sec61p. В некоторых случаях, однако, ретроградная транслокация не зависит от Sec61, и поэтому возникает вопрос о природе канала или даже о наличии нескольких каналов, специфичных для различных типов белков, обладающих неправильной структурой. Независимо от природы канала, для ретроградной транслокации надо выполнять те же шаги, как и для прямой транслокации.
Для того чтобы прошел экспорт, белок должен вначале попасть в канал, канал должен открыться, и затем — белок пройти через него в цитозоль.
Большинство белков направляются на деградацию только после завершения транслокации. Неизвестно, остаются ли эти белки поблизости от транслокона, пока не разрешится вопрос об их деградации. Также неизвестно, каким образом неправильно скрученные белки попадают в канал ретроградной транслокации. Фактически вырисовывающаяся картина позволяет предполагать, что путь, по которому направляется субстрат из ЭПР, и белки цитозоля, необходимые для деградации, сильно зависят от типа деградируемого белка и от того, где в нем располагается неправильно скрученная область.
Не исключено, что растворимые и трансмембранные белки деградируют по крайней мере частично отличающимся друг от друга путями и что среди трансмембранных белков эти пути зависят от положения мутации в домене люмена или цитозоля.
Как только белок оказался в ретроградном транслоконе, он, вероятно, начинает проходить через канал под действием сил цитозоля. Убиквитинилирование играет роль в экспорте многих (хотя и не всех) субстратов. Большинство субстратов деградации подвергается полиубиквитинилированию, и убиквитинилирование происходит, пока они еще находятся в связанном с мембраной состоянии. Мутации в каком-либо компоненте системы убиквитина приводят к образованию в ЭПР агрегатов, состоящих из белков неправильной структуры. Это позволяет предполагать, что канал блокируется в случае, когда субстрат не может модифицироваться.
Однако одного убиквитинилирования недостаточно для перехода белков в цитозоль. Также необходимо присутствие АТФазы цитозоля (р97 Cdc48 у дрожжей), которая связывается с мембраной ЭПР, при этом р97 связывает цитозольные кофакторы, которые прямо соединяются с субстратом ERAD. Пока неясно, каким образом эта АТФаза приводит к высвобождению субстрата в цитозоль, но аналогичные АТФазы бактерий и митохондрий непосредственно связываются с интегральными мембранными белками и извлекают их из мембраны. Не исключено, что АТФаза, требующаяся для ретротранслокации, действует аналогичным образом, связываясь или непосредственно с субстратом, или с присоединенным к нему остатком убиквитина и вытягивая белок из канала в цитозоль. Пока неизвестно, какую роль играет протеосома в процессе ретроградной транслокации.
Еще одним ключевым компонентом системы ERAD является небольшой мембранный белок дерлин. Этот белок необходим для ретроградной транслокации некоторых белков, обладающих неправильной структурой. Показано, что он взаимодействует как с неправильно скрученным белком в ЭПР, так и с р97 через связывание с белком VIMP. Поэтому дерлин представляет собой, по крайней мере, один компонент молекулярного «моста» между люменом ЭПР и аппаратом деградации цитозоля, хотя, конечно, существуют и другие.
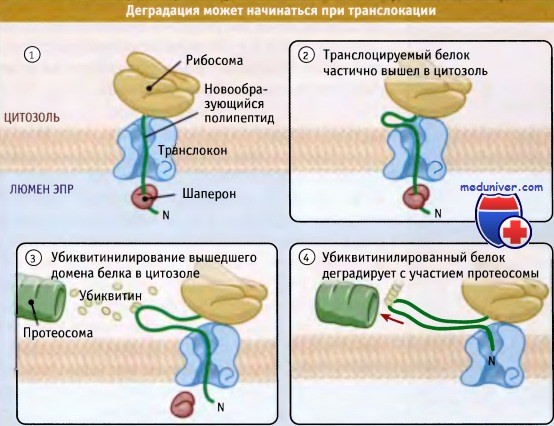
Как иллюстрирует рисунок ниже, в некоторых случаях белки могут отбираться для деградации даже до завершения их транслокации в ЭПР. Это используется для особенно больших субстратов для того, чтобы избежать затрат энергии на транслокацию белка целиком, если он начинает принимать неправильную конфигурацию уже в самом начале синтеза. Отбор белков для ретроградной транслокации в процессе их импорта позволяет не помечать их повторно. Наилучшим примером этому является деградация аполипопротеина В. Это очень большой секретируемый белок, который на первом шаге сборки липопротеинов низкой плотности связывается с липидами люмена ЭПР и жирными кислотами.
Неспособность фермента люмена ЭПР переносить липиды на аполипопротеин В по мере транслокации вызывает начало деградации белка до момента завершения его синтеза и транслокации.
Когда белки достигают цитозоля, они обычно узнаются и деградируют. Однако в некоторых случаях экспортируемые белки подвергаются агрегации в цитоплазме. При этом белки накапливаются в цитоплазме в специальных образованиях, которые называются агресомы. Они способны сохранять для деградации большие количества белка.
Исследование ERAD еще только начинается. Очевидно, в этом процессе участвует больше компонентов, чем известно на сегодняшний день, и его молекулярные основы исследованы пока недостаточно. Сюда относятся фундаментальные вопросы, связанные с распознаванием белков, обладающих неправильной структурой. Выяснение деталей этих процессов позволит реконструировать их in vitro.
Деградация некоторых белков начинается до того момента, как они полностью транслоцировались.
Когда белок отобран для деградации, в цитоплазме начинают накапливаться и убиквитироваться фрагменты его цепи.