• Веретено представляет собой комплекс, состоящий из микротрубочек и связанных с ними моторных белков. Организация микротрубочек обладает высоким уровнем поляризации
• Микротрубочки веретена представляют собой очень динамичную структуру. Одни проявляют динамическую нестабильность, для других характерна текучесть субъединиц
• Сила, необходимая для сборки веретена, генерируется при взаимодействии микротрубочек с моторными белками
Образование и функционирование веретена зависят от динамических свойств микротрубочек и от работы связанных с ними белковых моторов. Хотя микротрубочки образуют основные структурные элементы веретена, их организация и движение хромосом обеспечиваются белковыми моторами. Одни моторы непосредственно участвуют в сборке веретена и в связывании его компонентов в определенную структуру, а другие обеспечивают присоединение хромосом к веретену и генерируют силу, необходимую для их перемещения.
Несмотря на то что традиционно веретено рассматривается как структура, состоящая из микротрубочек, правильнее считать ее комплексом микротрубочек, белковых моторов и других белков.
Хотя моторы играют существенную роль в генерации силы, необходимой для функционирования веретена, микротрубочки представляют собой нечто большее, чем просто неподвижную структуру, вдоль которой они движутся. Во время митоза микротрубочки ведут себя как высокодинамичная структура, и это их свойство играет важную роль при сборке веретена и расхождении хромосом.
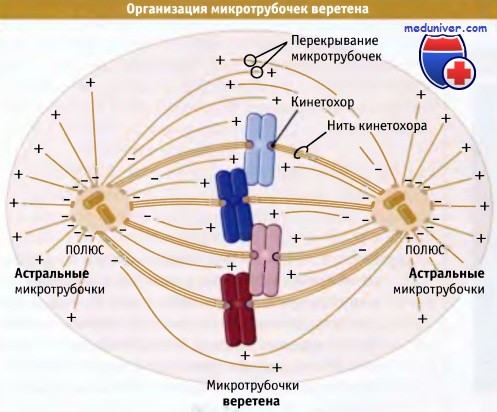
В веретене микротрубочки организованы в соответствии со своей полярностью.
Все минус-концы локализованы, поблизости от одной из двух центросом, а плюс-концы расположены на расстоянии от них.
В центре веретена микротрубочки от двух центросом перекрываются,
что обеспечивает расположение микротрубочек противоположной полярности (антипараллельные микротрубочки) близко друг к другу.
В пределах веретена микротрубочки организованы в соответствии с полярностью. Два конца микротрубочки различаются по составу и структуре. Это обусловливает ее структурную «полярность»; микротрубочка как бы указывает то или иное направление. В каждом полуверетене и связанной с ним звезде микротрубочки расположены с одинаковой полярностью: их минус-концы находятся на полюсах, а плюс-концы, на некотором от них расстояниии.
В месте пересечения двух поляризованных пучков микротрубочки перекрываются, создавая область в центре веретена, в которой соседние микротрубочки имеют противоположную полярность. Одинаковая ориентация микротрубочек в каждом полуверетене необходима для нормального функционирования их моторов при делении. Если бы полярность микротрубочек в пределах каждого полуверетена была произвольной, то молекулы каждого типа моторов просто мешали бы друг другу, делая движение хаотичным или просто невозможным.
Динамические свойства микротрубочек играют важную роль во всех фазах митоза. Исследования, проведенные на культуре клеток позвоночных и с использованием экстрактов из яйцеклеток лягушки Xenopus laevis, показали, что в каждом веретене микротрубочки характеризуются динамической нестабильностью и являются более короткими и гораздо более динамичными, чем в интерфазных клетках. Некоторые различия можно объяснить возрастанием частоты катастроф в митозе, когда плюс-концы микротрубочек из состояния роста или полимеризации переходят в состояние укорочения или разрушения. Частично это также объясняется снижением частоты наступления спасений, при которых процесс деполимеризации или укорочения микротрубочек обратно переходит в процесс их полимеризации или роста.
Это усиление динамики происходит в клетках, вступающих в митоз, поскольку белки, связанные с микротрубочками и обычно препятствующие катастрофе, заингибированы, в то время как другие, стимулирующие рост микротрубочек, активируются. Баланс между двумя противоположно направленными процессами поддерживается основной киназой, регулирующей митоз, комплексом циклин B/CDK1, которая активируется во время разрушения ядерной оболочки. Как будет показано ниже, усиление динамики микротрубочек в клетках, вступающих в митоз, играет основную роль в сборке веретена.
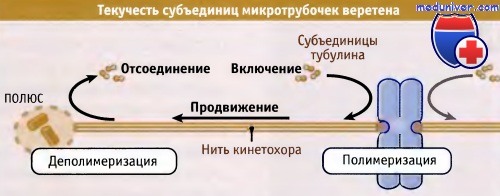
После образования веретена начинает проявляться еще один тип динамики микротрубочек. В это время микротрубочки обнаруживают текучесть субъединиц. Это интересное явление заключается в том, что субъединицы тубулина присоединяются к плюс-концу микротрубочки и затем продвигаются по ней к минус-концу, на котором высвобождаются. Как следует из рисунков ниже, текучесть характерна для всех микротрубочек веретена, однако особенно она проявляется у микротрубочек нитей кинетохора. Происхождение этого явления не вполне понятно, но, возможно, оно связано с взаимодействием плюс- и минус-концов микротрубочек веретена с другими его компонентами (например, с белковыми моторами). Даже в то время, когда у микротрубочек веретена наблюдается текучесть, астральные микротрубочки продолжают проявлять динамическую нестабильность.
Хотя значение явления текучести неизвестно, возможно, оно играет роль в перемещении хромосом и в поддержании баланса сил в веретене, с тем чтобы две его половины оставались расположенными симметрично.
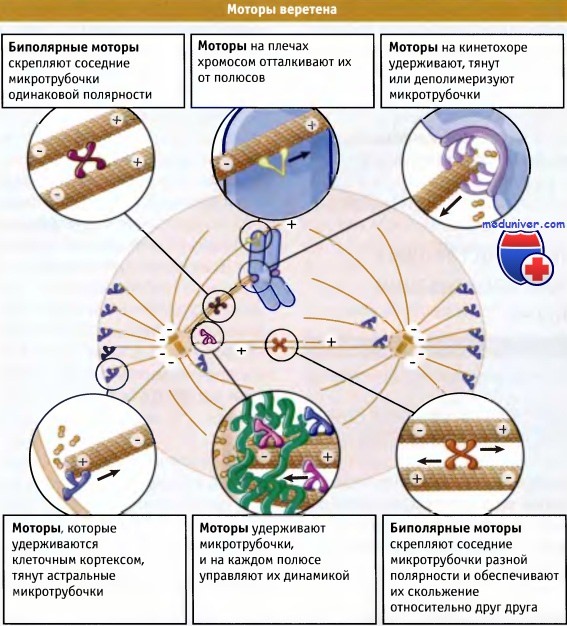
С системой микротрубочек взаимодействуют много различных типов белковых моторов. В митозе участвует цитоплазматический мотор динеин, осуществляющий транспорт к минус-концу, и моторы группы кинезинов (большая часть которых движется в направлении плюс-конца). Веретено имеет сложную организацию, и моторы настолько тесно связаны с его формированием и функцией, что только в делении клеток высших организмов участвует более 15 представителей семейства кинезинов.
Белковые моторы расположены по всему веретену. Они находятся на кинетохорах, на плече хромосом, на полюсах и на микротрубочках между полюсами и хромосомами. Многие типы моторов располагаются только в определенных местах, другие занимают несколько мест. Например, цитоплазматический динеин обнаружен в кинетохорах и на полюсах, а также в клеточном кортексе, где он взаимодействует с астральными микротрубочками. В то же время кинезин-подобный белковый мотор CENP-E находится в кинетохоре, а хромокинезины только на плечах хромосом.
В митозе белковые моторы выполняют несколько основных функций. Одни из них, например динеин, связываются со структурами, включая кинетохоры и плазматическую мембрану, и транспортируют их вдоль микротрубочки (хотя в случае плазматической мембраны движется микротрубочка). Другие имеют множественные домены, организованные таким образом, что мотор может связываться сразу с двумя микротрубочками, и скреплять их между собой. В зависимости от структуры моторов микротрубочки в пучке могут обладать той же самой или противоположной полярностью. Если мотор связывается с микротрубочками противоположной полярности, он будет пытаться двигаться (скользить) по ним до тех пор, пока они перекрываются. Примером такого типа моторов является представитель кинезинов Eg5, который может связываться с обоими концами антипараллельных микротрубочек.
Наоборот, если мотор устроен так, что он связан с двумя микротрубочками с одинаковой полярностью, то в результате образуется структура с такой же полярностью, расположенная таким образом, что микротрубочки образуют фигуру, напоминающую звезду. Прочие кинезин-подобные белки не перемещаются по микротрубочкам, а способствуют разборке их плюс-концов. Наглядным примером такого белка является кинезин, связанный с митотической центромерой (МСАК), который находится на центромере каждой хромосомы. В состав веретена входят моторы с перечисленными выше основными свойствами, которые определенным образом расположены относительно друг друга. Эти же моторы генерируют усилия для движения хромосом.
Не всегда ясно, каким образом моторы обеспечивают функционирование веретена. В ряде случаев, например, они располагаются таким образом, что могут мешать друг другу. Однако, независимо от деталей строения веретена, очевидно, что его образования и функционирования необходимы множественные сбалансированные усилия. Эти усилия обеспечиваются моторами, которые расположены на каркасе динамических микротрубочек веретена.
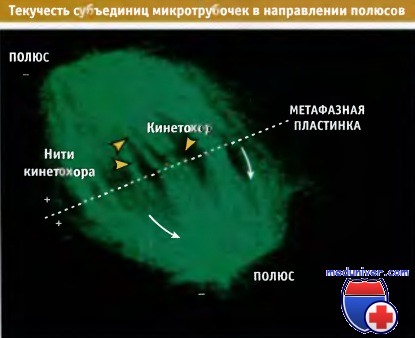
Субъединицы тубулина постоянно включаются в микротрубочки со стороны кинетохоров и продвигаются к полюсам, где происходит их высвобождение.
Таким образом, они постоянно мигрируют от кинетохоров к полюсам вдоль микротрубочек нити кинетохора.
В течение метафазы длина кинетохорной микротрубочки остается постоянной, пока скорость сборки субъединиц на плюс-конце соответствует их разборке на минус-конце.
Если сборка субъединиц со стороны кинетохора снижается, а на полюсе скорость их разборки не изменяется, то кинетохор будет двигаться к полюсу.
Таким образом, текучесть субъединиц микротрубочек представляет собой возможный способ движения хромосомы.
Первый видеокадр, на котором представлено митотическое веретено клетки, часть тубулина которого содержит флуоресцирующий зонд (флуоресцирует зеленым).
Кинетохоры выделены оранжевыми стрелками. На видео показан поток зеленых точек кинетохорной нити во всем веретене.
В образовании веретена участвуют молекулярные моторы, которые перемещаются по микротрубочкам.
Веретено формируется за счет специфических взаимодействий между этими моторами и микротрубочками.
Эти взаимодействия обеспечивают также его подвижность и являются источниками силы.
Стрелками указано направление движения моторов.