Генетика острого промиелоцитарного лейкоза при транслокации t(15;17)(q22;ql2-21)
Как отмечалось, для острого промиелоцтарного лейкоза характерна t(15;17)(q22;ql2-21). В результате этой транслокации так называемый PML-ген (ген промиелоцитарного лейкоза), расположенный на хромосоме 15, переносится на длинное плечо хромосомы 17 в область, где находится ген а-рецептора ретиноевой кислоты (RARa).
Ген RARa (рецептора ретиноевой кислоты а) относится к семейству рецепторных генов (гены рецепторов стероидных гормонов, эстрогенов, тиреоидных, витамина D3), являющихся транскрипционными факторами, которые в присутствии определенных лигандов способны либо активировать, либо подавлять трансактивацию необходимых генов. Лигандом для гена RARa является ретиноевая кислота. В норме этот ген участвует в регуляции дифференцировки миелоидных клеток.
Давно отмечено, что ретиноиды играют ключевую роль в миелоидной дифференцировке, поскольку при дефиците витамина А и у человека, и у экспериментальных животных отмечаются нарушения гемопоэза, при этом введение ретиноидов стимулирует преимущественно гранулоцитопоэз. Ретиноиды (полностью транс-ретиноевая кислота, 13-цис-ретиноевая кислота, 9-цис-ретиноевая кислота и др.) являются лигандами ядерного рецептора RARa, который прикрепляется к ДНК в регионах связывания ретиноевой кислоты (RARE) только после гетеродимеризации с другим ретиноевым рецептором X (RXR).
В отсутствие лигандов (ретиноидов) гетеродимер связывается с корепрессорами SMRT и N.COR, которые в свою очередь связаны с гистондеацетилазным-Sin3A комплексом, что ведет к репрессии транскрипции необходимых генов. Гистондеацетилазы ингибируют механизмы транскрипции за счет прикрепления молекулы ДНК к гистонам (compact DNA on histones). При связывании ретиноевой кислоты с RARa происходит обмен корепрессоров на коактиваторы транскрипции, которые связаны с гистонацетилазами, что приводит к откреплению ДНК от гистонов и активации транскрипции необходимых генов.
К генам, регулируемым RARa, относятся гены регуляторов клеточного цикла (циклины, циклинзависимые киназы), молекул адгезии (CD11b, CD18), интерлейкинов, моноцитарного хемоаттрактанта, колониестимулирующих факторов (Г-КСФ, ИЛ-1, ИЛ-8), рецепторов колониестимулирующих факторов (рецепторы моноцитарного КСФ и Г-КСФ), регуляторов апоптоза и терминального клеточного деления (трансглутаминаза II, bс12), свертывающих факторов (тромбомодулин, тканевый фактор, урокиназа, тканевый активатор плазминогена и их ингибиторы), гены факторов транскрипции (STAT, НОХ, другие RAR).
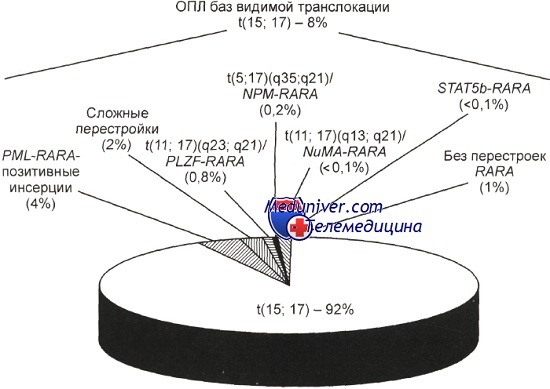
Гистограмма, отражающая частоту специфических хромосомных и молекулярно-генетических маркеров при остром промиелоцитарном лейкозе
Ген PML считается геном-регулятором роста и играет роль в созревании и активации различных клеток. Продуктом этого гена является туморсупрессирующий протеин, участвующий в процессах дифференцировки клеток и подавления их пролиферации, в ряде иммунологических процессов, связанных, в частности, с механизмами действия ИФН-a. Так, показано, что PML-протеин стимулирует экспрессию антигенов I класса главного комплекса гистосовместимости и белков, участвующих в передвижении пептидов к поверхности клеток в ассоциации с антигенами I класса. В ряде работ было продемонстрировано, что PML-протеин может индуцировать процесс апоптоза, как связанный, так и не связанный с каспазным механизмом. Белок PML экспрессируется в основном в дифференцированных клетках в постмитотическом периоде.
Самая большая экспрессия этого белка обнаруживается в клетках эндотелия, эпителиальных клетках и макрофагах. Белок PML локализуется в ядре клетки в так называемых ядерных тельцах (nuclear bodies — NB) или PML-онкогенных доменах (POD). Эти структуры были описаны около 35 лет назад, их присутствие прямо пропорционально уровню белкового синтеза и обратно пропорционально степени дифференцировки клеток. NB связаны с ядерным матриксом, который играет роль в перемещении молекул и организации хроматина внутри ядра. При остром промиелоцитарного лейкоза с t(15;17) белок PML перемещается из NB и визуализируется как мелкодисперсный материал. После лечения полностью транс-ретиноевой кислотой PML вновь локализуется в ядерных тельцах. Исследователи отметили, что увеличение количества белка PML в клетках культуры острого промиелоцитарного лейкоза (NВ4-линия) существенно подавляет ее клоногенную активность и злокачественность при проведении экспериментов на бестимусных мышах (nude). Это позволило авторам предположить антагонизм между действием белка PML и белкового продукта химерного гена PML-RARa.
Продуктом химерного гена PML-RARa является патологический белок, который сохраняет в себе активные функциональные домены как белка RARa, так и белка PML. При ОПЛ белок PML-RARa накапливается в цитоплазме и ядре миелоидных клеток в существенно большем количестве, чем в норме накапливается белок RARa. Аберрантный ретиноидный рецептор PML-RARa с нарушенной ДНК-связывающей активностью может прикрепляться к ДНК в регионах связывания ретиноевой кислоты (RARE) как гомодимер, конкурируя с нормальным RARa, который может связываться с ДНК, как указывалось, только после гетеродимеризации с RXR.
Также доказано, что химерный белок активно связывается и с RXR, вытесняя нормальный рецептор RARa. В отсутствие ретиноевой кислоты химерный рецептор PML-RARa оказывается более сильным репрессором транскрипции, чем нормальный рецептор. Это объясняется тем, что, формируя репрессор-ный белковый комплекс, он сильнее, чем нормальный RARa, связывается с корепрессорными молекулами N.COR и SMRT. Эти корепрессорные молекулы в свою очередь связаны с гистодеацетилазами, которые изменяют конформацию молекулы ДНК и делают ее недоступной для транскрипционных факторов. Вследствие этого транскрипция генов останавливается. Чтобы вызвать диссоциацию комплекса RARa-корепрессоры-гистондеацетилазы, концентрация ATRA должна составлять 10-6 моль/л.
Это значительно превышает физиологическую концентрацию ретиноевой кислоты (10-9 моль/л), которая требуется для диссоциации комплекса, в состав которого входит нормальный рецептор RARa. В норме после связывания лиганда (ATRA) с лигандсвязывающим доменом RARa происходит отсоединение корепрессоров (диссоциация комплекса RARa-корепрессоры-гистондеацетилазы), меняется конфигурация рецептора RARa, в результате чего появляются домены ассоциации с коактиваторами транскрипции TIF1/TIF2/CBP. При ОПЛ в условиях низкой физиологической концентрации ретиноевой кислоты химерный белок PML-RARa удерживает корепрессордеацетилазный комплекс, что приводит к замедлению процесса активации транскрипции и блокирует транскрипцию генов миелоидной дифференцировки. Этот блок дифференцировки может быть снят только при высокой концентрации ретиноевой кислоты, что достигается во время терапии полностью транс-ретиноевой кислотой (ATRA).
Эффекты белка PML-RARa связаны не только с блоком дифференцировки, но и с регуляцией апоптоза и роста клеток. Так, in vitro клетки, экспрессирующие указанный белок, не подвергаются апоптозу в тех ситуациях, когда удаляются факторы, необходимые для поддержания их жизнеспособности (сыворотка или гранулоцитарно-макрофагальный колониестимулирующий фактор — Г-КСФ), тогда как в контроле (в случае, если отсутствует экспрессия PML-RARa) клетки погибают. Из этого следует, что этот белок поддерживает жизнеспособность опухолевых клеток, блокируя механизмы апоптоза.