Влияние продуктов питания на полярность эпителия кишечника
Деятельность клеток скелетных мышц не зависит от полярности клеток. Например, мышечные клетки получают сигналы о поступлении глюкозы от любой точки цитоплазматической мембраны, и, насколько можно в настоящее время предполагать, изменения в экспрессии генов не зависят от места попадания глюкозы в клетку. Клетки в составе эпителия различаются по своей полярности. Это разделение между апикальной (граничащей с содержимым кишки в случае кишечного эпителия) и базолатеральной частями является существенным для активности эпителия. Данное свойство клеток хорошо изучено относительно переноса нутриентов через эпителий кишки. Существуют различия в переносе через апикальную и базолатеральную мембраны ионов, молекул малого размера и макромолекул.
Именно полярность эпителия определяет направление движения этих субстанций через эпителий (внутрь клетки или наружу из нее). Данное свойство следует учитывать также и при обсуждении других функций эпителиальной клетки. Полярность особенно важна при исследовании взаимодействия нутриентов и генов в кишечном эпителии. Полярность эпителиальных клеток характеризует два важнейших механизма, посредством которых нутриенты (и другие компоненты содержимого кишки) оказывают влияние на гены:
(1) прямое воздействие содержимого кишки на энтероциты;
(2) непрямое воздействие, осуществляемое через гормоны, фактор роста и цитокины.
Более того, кишечный эпителий выступает в роли барьера между организмом и внешней средой, которую в данном случае представляет содержимое кишки. Этот барьер не является полностью непроницаемым, поскольку кишечник выборочно «позволяет» макромолекулам проходить через него. Кроме того, происходит активная абсорбция нутриентов. Постепенно стало понятно, что энтероцит выступает в роли иммунной клетки. Он имеет рецепторы для продуктов деятельности бактериальных клеток, а также участвует в экспрессии разнообразных молекул на своей поверхности, имеющей иммуноглобулиновые домены. Эпителиальная клетка также экспрессирует белки, которые могут взаимодействовать с иммуноцитами в кишечнике. К этим белкам относят такие поверхностные молекулы, как молекулы главного комплекса гистосовместимости (ГКГС) класса II и те цитокины, которые высвобождаются из эпителия, например хемокины или интерлейкин (IL) 6.
Перечисленные сигнальные белки дают возможность эпителиальной клетке играть ведущую роль в процессах, происходящих в кишечнике. Исследовательская группа выдвинула гипотезу, что изменения содержимого кишечника регулируют экспрессию эпителиальной клеткой сигнальных молекул. Это означает, что изменения содержимого кишечника в результате вариаций в диете оказывают влияние через эпителий кишечника на другие его структуры, особенно на иммунную систему его слизистой оболочки. Существуют два компонента сигнального пути, связывающие питание и бактерии, находящиеся в просвете кишки, с иммунной системой слизистой оболочки:
(1) афферентный путь, включающий механизмы, посредством которых изменение кишечного содержимого влияет на экспрессию генов в кишечном эпителии;
(2) эфферентный путь, представляющий собой действие белков, экспрессированных эпителиальной клеткой, действующей как компонент иммунной системы кишечника. В настоящее время существуют доказательства функционирования обоих путей.
Современная наука позволяет изучить изменения, происходящие с молекулами в эпителии кишки в ответ на изменения в диете (афферентный путь). В качестве экспериментальной модели может выступить экспрессия молекул ГКГС класса II в эпителии мыши, которая только что перестала получать грудное молоко. Молекулы ГКГС класса II ответственны за презентацию антигена, их экспрессия на эпителии кишечника мышей отмечается после их отлучения от груди, поэтому возможен перевод мышей на привычную для них пищу (корм для мышей) или определенного состава жидкую смесь (энтеральное питание) для изучения разницы между двумя видами питания. Эпителиальные клетки мыши исследовали в разные моменты времени после прекращения кормления ее грудным молоком. При этом изучали экспрессию молекул ГКГС класса II и инвариантную цепь, которая также подвергается экспрессии.
Воздействие энтерального питания (Vivonex) значительно отличалось от такового при кормлении мышей обычным для них кормом.
В норме корм для мышей индуцирует экспрессию генов сигнальных молекул между 20 и 30 днями жизни, в то время как энтеральное питание за этот же период не приводит к экспрессии генов в кишечном эпителии. В настоящее время доказано, что регуляторный нуклеарный белок (трансактиватор класса II, СIIТА) является необходимым и достаточным условием для экспрессии молекул ГКГС класса II.
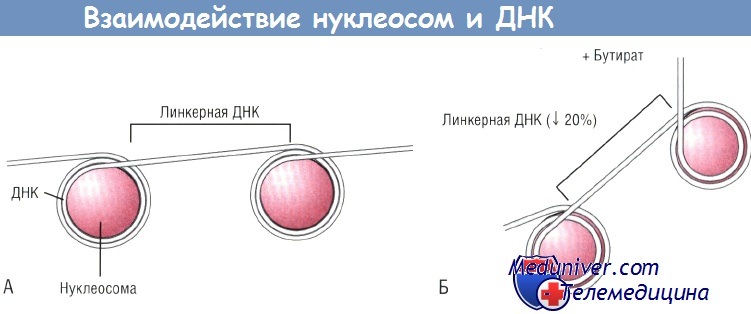
(А) ДНК делает два полных оборота вокруг нуклеосом, состоящих из неацетилированных гистонов.
(Б) При бутират-индуцированном ацетилировании нуклеосомы раширяются, при этом уменьшается количество оборотов ДНК вокруг нуклеосом до 1,8 и сокращается длина связующего их участка ДНК (линкерной ДНК).
В результате ДНК не может линейно следовать от нуклеосомы к нуклеосоме, поворачивается под углом после каждой нуклеосомы, что ведет к нарушению совокупности нуклеосом. ДНК — дезоксирибонуклеиновая кислота.
Были проведены эксперименты с целью изучить, действительно ли влияние питания опосредовано трансактиватором класса II. У мышей были выявлены три изоформы СИТА. Интересно, что нормальная комплексная диета повышала экспрессию молекул ГКГС класса II посредством СIIТА IV. Кроме регуляции молекул ГКГС класса II посредством диеты отмечалась еще и медленная, зависимая от времени регуляция посредством СIIТАIII. Данные эксперименты показывают, что изменения в диете соответствуют определенным изменениям молекулярных взаимосвязей между содержимым кишечника и механизмом сигнальной трансдукции в эпителиальной клетке.
Бактериальная ферментация тех углеводов, которые не абсорбировались в кишечнике, приводит к продукции короткоцепочечных жирных кислот (КЦЖК). Это означает, что уровень бутирата отражает изменения в популяции бактерий и в тех субстратах, которые доступны для метаболизма бактерий. Уровень бутирата сильно зависит от внешних воздействий. Например, у новорожденных детей отмечают очень низкие уровни бутирата как в тонкой, так и в толстой кишке; уровень бутирата в организме ребенка достигает уровня взрослого к 2-летнему возрасту. Интересно, что в течение первых 6 мес жизни уровни бутирата намного выше у детей, находящихся на искусственном вскармливании, чем у детей, получающих грудное молоко.
Следовательно, уровень бутирата отражает те изменения, которые происходят в содержимом кишечника. Было высказано предположение, что концентрация бутирата может изменять активность сигнала в эпителиальной клетке. Исходя из этой гипотезы было исследовано воздействие бутирата на IL-8 и экспрессию моноцитарного хемотаксического протеина 1 (МСР-1). Увеличение концентрации бутирата повышало секрецию IL-8 при одновременном снижении экспрессии МСР-1. Эти эффекты были обнаружены в популяциях неактивных эпителиальных клеток, но наибольшие изменения были характерны для клеток, стимулированных такими провоспалительными агентами, как липополисахариды (ЛПС) или IL-1b.
Известно, что бутират изменяет ацетилирование гистонов. Нуклеосома состоит из соленоида гистонов, вокруг которого дважды «обернута» ДНК. Бутират повышает ацетилирование гистона, тем самым снижая его компактность. В результате ДНК не может «обернуться» вокруг большой нуклеосомы полным оборотом, а нуклеосома теряет способность упаковываться в плотную структуру. Это «обнажает» ДНК и делает ее более подверженной действию факторов транскрипции. Было высказано предположение, что таким образом бутират изменяет экспрессию хемокинов. Для проверки данной гипотезы был использован фунгицид трихостатин A (TSA), в 700 раз более мощный индуктор ацетилирования гистонов по сравнению с бутиратом.
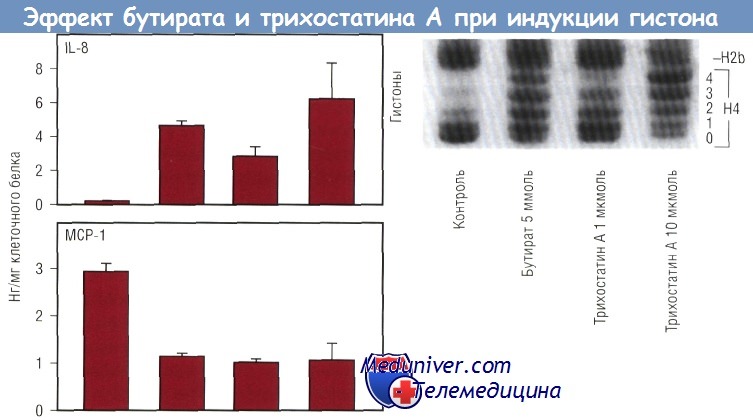
Если бы действие бутирата на секрецию хемокина было обусловлено повышением ацетилирования гистонов, то можно было бы ожидать подобного эффекта и от TSA. Опыты с TSA показали, что TSA повышал секрецию IL-8 и снижал секрецию МСР-1. В нижней части рисунка продемонстрировано, что и TSA, и бутират повышали ацетилирование гистона-4. Неацетилированные гистоны быстрее продвигались через гель и образовывали одну полоску («ступеньку»), в то время как ацетилированные гистоны образовывали «лестницу» в зависимости от степени ацетилирования. Гистон-4 имеет 4 ацетилированных остатка лизина. Таким образом, ацетилирование гистонов будет отражено в образовании «лестницы» из 5 «ступенек». Это можно увидеть при действии бутирата или TSA. Верхняя часть рисунка показывает, что TSA дает такой же эффект, как и бутират. TSA повышает секрецию IL-8 и снижает секрецию МСР-1. Более того, степень увеличения IL-8 меняется в зависимости от степени ацетилирования гистонов. Эффект бутирата на ацетилирование гистонов обратим.
Ацетилирование и секреция хемокина клетками Сасо-2. Как бутират, так и трихостатин А увеличивали ацетилирование гистонов. Одновременно увеличивалась секреция IL-8 и уменьшалась продукция МСР-1 в клетках Сасо-2, стимулированных IL-1.
Трихостатин А, специфический ингибитор гистондеацетилазы, действовал так же, как и бутират, когда поступал в концентрациях, приводящих к сравнимому изменению ацетилирования гистона.
На диаграмме представлены стандартные отклонения трех различных ячеек для каждого вещества, отражающие результаты трех экспериментов. IL — интерлейкин; МСР — моноцитарный хемотаксический протеин.
После прекращения действия бутирата ацетилирование гистонов возвращается к норме и отмечается такое же действие IL-1b на секрецию IL-8 и секрецию МСР-1, как до эксперимента. КЦЖК с различной длиной цепи оказывают дифференцированное влияние на ацетилирование гистонов. Бутират является наиболее действенной КЦЖК, индуцирующей ацетилирование гистонов, в то время как от кислот с большей или меньшей углеводной цепью наблюдаются менее выраженные эффекты. Данное влияние на ацетилирование гистонов обусловлено действием на экспрессию IL-8 и МСР-1. Из всех использованных в опыте КЦЖК именно бутират вызывает наибольшее увеличение секреции IL-8 и наиболее выраженное уменьшение секреции МСР-1.
Итак, эти опыты показывают, что бутират изменяет экспрессию хемокинов в эпителиальной клетке. КЦЖК изменяют экспрессию посредством ацетилирования гистонов. Данные эксперименты, однако, не исключают возможность того, что дополнительное действие бутирата может быть опосредовано промоторными системами. Действительно, недавно проведенные исследования показали, что бутират снижает экспрессию связывающего инсулиноподобный фактор роста белка (IGFBP) 3, посредством ацетилирования протеина, ингибирующего связывание ДНК. В дальнейшем необходимо исследовать взаимосвязь между хромосомной регуляцией, продемонстрированной в приведенном эксперименте, и регуляцией, обусловленной промотором, как для бутирата, так и для других молекул содержимого кишечника.
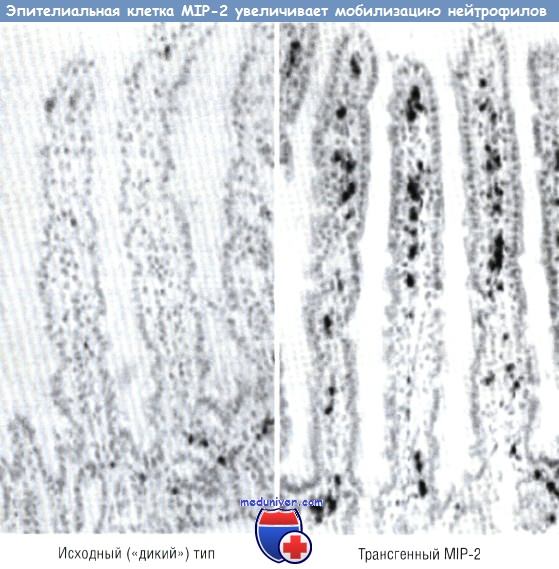
Доказательство действия экспрессии генов эпителиальных клеток на иммунную систему слизистой оболочки (эфферентного пути) основано на возможности селективного изменения экспрессии генов в эпителиальной клетке кишечника посредством трансгенных технологий. В качестве модели для демонстрации управления иммунной системой слизистой оболочки эпителием была использована экспрессия хемокинов в эпителии. Хемокин IL-8, который у человека отвечает за мобилизацию нейтрофилов, был первым из идентифицированных хемотаксических цитокинов. Однако IL-8 у мышей не экспрессирован. Для изучения действия хемокинов на иммунную систему слизистой оболочки была разработана система, с помощью которой макрофагальный воспалительный белок 2, имеющий эффект, практически подобный эффекту IL-8 у человека, был присоединен к промотору FABPI (белок кишечника, связывающий жирную кислоту).
Промотор активен только в эпителиальных клетках тонкой кишки и проксимального отдела толстой кишки. Была разработана специальная «конструкция», где промотор FABPI и макрофагальный воспалительный белок 2 комплементарной ДНК (кДНК) были присоединены к интрону и полиадениловому «хвосту». Эта «конструкция» была внедрена в яйцеклетку мыши. Было выявлено, что эпителий первой генерации экспрессирует макрофагальный воспалительный белок 2 мРНК. Анализы показали воздействие этой «конструкции» на мобилизацию как нейтрофилов, так и лимфоцитов. У трансгенной мыши отмечали повышенную мобилизацию нейтрофилов в собственной пластинке слизистой оболочки кишечника и в эпителиальных клетках. Эффект хемокина был отмечен только в тех клетках, где промотор FABPI был активен. В тонкой кишке, где промотор FABPI активен, мобилизация нейтрофилов, выражающаяся активностью миелопероксидазы (на единицу массы кишки), была значительно выше у трансгенных мышей. В проксимальной кишке, где промотор FABPI также активен, отмечали увеличение нейтрофильной инфильтрации.
Однако в дистальном отделе толстой кишки, где промотор FABPI неактивен, подобных эффектов отмечено не было. Кроме того, в печени и селезенке трансгенных мышей не выявляли увеличения инфильтрации больше, чем у нормальных мышей (в этих органах промотор FABPI неактивен). На первый взгляд, эти данные свидетельствуют о том, что эпителиальная клетка может посредством высвобождения хемокинов изменять иммунную функцию слизистой оболочки в кишечнике in vivo.
У трансгенной мыши MIP-2 экспрессирует эпителий кишечника, используя эпителий-специфический промотор. Нейтрофилы были локализованы с помощью миелопероксидазы.
MIP — макрофагальный воспалительный белок
Дальнейший анализ иммунной системы показал, что в тонкой кишке помимо инфильтрации нейтрофилов была отмечена повышенная инфильтрация лимфоцитов. Их количество в собственной пластинке слизистой оболочки было увеличено. Также было отмечено удвоение числа лимфоцитов в эпителиальном слое слизистой оболочки кишки. Исследование рецепторов, расположенных на поверхности лимфоцитов, локализованных в эпителии, показало, что лимфоциты экспрессировали CXCR2, который является рецептором, отвечающим за трансдукцию активности макрофагального воспалительного белка 2. Таким образом, эксперименты показывают, что изменение экспрессии только одного хемокина в эпителии влияет на функцию как лимфоцитов, так и нейтрофилов.
Однако изменения содержимого кишечника могут оказывать воздействие на многие хемокины. Кроме того, вероятен эффект воздействия на иммунную систему и других цитокинов. Становится понятно, что изменения экспрессии генов в эпителии оказывают большое влияние на остальные составляющие иммунной системы слизистой оболочки.
Существует гипотеза, что эти сигнальные процессы важны не только для понимания физиологии здорового организма, но и для лечения заболеваний. Например, начальным этапом терапии детей с болезнью Крона в Великобритании является лечебное питание. Несмотря на то что известны многие механизмы, посредством которых энтеральное питание проявляет свои лечебные свойства, есть мнение, что еще одним методом может быть радикальная смена содержимого кишечника до такой степени, что изменятся сигналы, идущие от эпителия кишечника к иммунному компоненту слизистой оболочки, в результате снизится экспрессия воспаления при болезни Крона. Этим методом можно достигать уменьшения активности воспалительного процесса при данной патологии.