Методы оценки кинетики белков, аминокислот в организме
Описанные методы могут дать лишь общее представление о метаболизме белка в организме. Это упрощенный взгляд, поскольку метаболизм белка присущ разным тканям и идет он с разной степенью интенсивности. Например, некоторые белки (в частности, ферменты печени) имеют период полураспада, равный нескольким минутам, в то время как мышечный белок меняется не более чем на 1—2% в сутки.
Эта информация побуждает к поискам новых подходов к оценке скорости обмена конкретных белков или белков в разных тканях.
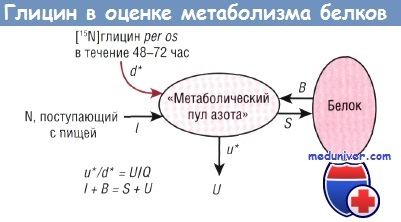
При инфузии любой меченой аминокислоты индикатор распределяется в пуле свободных аминокислот в большинстве тканей, часть индикатора участвует в синтезе белка и встраивается в структуру любого белка тела человека. Возможность провести измерение с помощью газовой хромато-комбустион-изотопной масс-спектрометрии относительно низкого изотопного обогащения специфических белков позволила определить скорость синтеза этих белков. Например, альбумин сыворотки можно получить из неоднократно взятых образцов плазмы в ходе инфузии [15N]глицина в течение 72 час.
Затем полученный белок подвергают гидролизу in vitro. Можно измерить увеличение 15N-обогащения связанных с альбумином остатков глицина. Фракцию альбумина, синтезированного за единицу времени, называют скоростью фракционного синтеза (%/сут). Расчет ее прост: необходимо разделить прирост обогащения связанного с альбумином [15N]глицина на прошедшее время и на обогащение в пуле предшественника свободного глицина. В качестве предшественника, используемого для расчета, берут как свободный глицин плазмы, так и гиппурат (другой продукт метаболизма глицина, предположительно образующийся из такого же внутриклеточного печеночного пула, но который можно выделить из мочи).
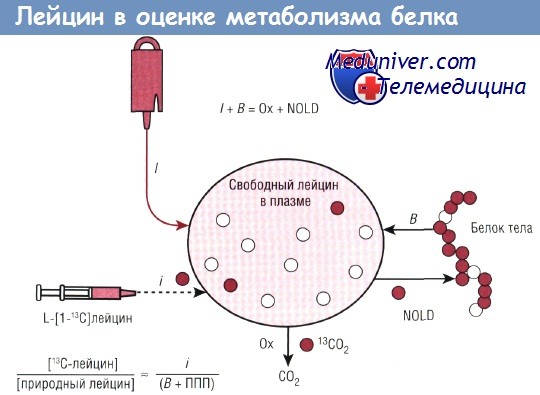
Кинетика заменимой аминокислоты также может быть определена путем комбинированного введения [13С]лейцина и меченого атома интересующей аминокислоты, например [15N]глютамина. Поскольку глютамин является заменимой аминокислотой, скорость его продукции (RaGln) в состоянии натощак обусловливают два процесса: высвобождение глютамина при распаде белка и новый синтез глютамина.
Учитывая, что в результате распада белка свободные аминокислоты вывобождаются пропорционально их относительному избытку в виде связанных остатков в белке организма, высвобождение глютамина в результате протеолиза можно оценить как RaLeu х 0,421, где RaLcu — постабсорбтивная скорость образования лейцина, а 0,421 — отношение глютамина к избытку лейцина, отражающее остатки в белке организма (единица измерения для обоих — мкмоль аминокислоты/г белка). Часть RaGln, которая не может быть определена при распаде белка, включается в новый синтез глютамина. Такой подход был использован для демонстрации способности организма детей с ОНМТ синтезировать глютамин de novo через 10 сут жизни.
Для выяснения, могут ли некоторые аминокислоты, относящиеся к заменимым у взрослых, быть незаменимыми у младенцев, Miller и соавт. применили следующий метод: они мониторировали появление 13С в заменимых аминокислотах после назначения [U-13С]глюкозы и наблюдали очень небольшое обогащение 13С в цистеине плазмы, что позволило сделать предположение об условной незаменимости цистеина у новорожденных. Для оценки регионарной кинетики аминокислот также использовали технологии двойного изотопа.
При комбинации внутривенной инфузии L-[13С5]глютамина с пероральным введением L-[1-13С5]глютамина было установлено, что 46—53% поступающего энтеральным путем глютамина выводится через внутренние органы (кишечник, печень), если основываться на обнаружении энтерально вводимого меченого атома в плазме. Эти данные позволяют предположить возможное значение глютамина как «энергетического топлива» в кишечнике новорожденного.
Итак, в течение двух последних десятилетий произошли существенные изменения в разработке неинвазивных технологий по исследованию питания и метаболизма у новорожденных детей. Значимость этих изменений можно сравнить со значимостью кинематографа, который пришел на смену длительно существовавшей статичной, обездвиженной фотографии.
Для исследователей стали доступны этически приемлемые методы оценки динамических потоков энергетических субстратов, белков и специфических нутриентов. Эти технологии позволили получить огромное количество информации о потоке нутриентов в организме новорожденного ребенка. Несмотря на то что большинство упомянутых технологий остаются ограниченными рамками исследовательских проектов, полученная информация уже успешно внедрена в реализуемые стратегии по питанию новорожденных, и этот процесс будет продолжаться.