Апоптоз и прекращение клеточного цикла в G1-фазе представляют собой два различных способа прекращения роста клетки, которые приводят к ее неспособности войти в S-фазу. Апоптоз является эволюционным механизмом, используемым всеми многоклеточными организмами в регуляции роста и дифференцировки клеток. Существуют два основных пути апоптоза: внутренний и внешний. Внутренний путь запускается несколькими внутриклеточными сигналами, генерируемыми в ответ на гипоксию, повреждение ДНК, cis- или trans-активацию онкогенеза, либо ограничивающим пороговым сигналом по ходу каскада факторов роста.
При внешнем пути (апоптозе, индуцируемом активацией) внеклеточные агенты запускают лишь частично изученный каскад внутриклеточных трансдьюсеров сигналов, которые сводят вместе биохимические сигналы и сигналы-регуляторы генов, вызывающие серию морфологических изменений. В итоге оба пути приводят к конденсации хроматина и расщеплению ДНК внутри нуклеосомы. В противоположность апоптозу клетки, у которых развитие прекратилось в G1-фазе клеточного цикла, сохраняют жизнеспособность и ДНК у них остается интактной.
Увеличиваются доказательства, позволяющие предположить, что в регулирование этих путей вовлечено некоторое число генов параллельно с р53, кальциневрин/кальмодулин и IGF/инсулин-индуцибельными сигнальными путями. Таким образом, цикл клетки, по всей видимости, обусловлен общим набором молекул-трансдукторов и последующим контролем клеточной пролиферации, прекращения роста или апопто-за, осуществляемым через генерирование или ингибирование сигналов выживания или смерти.
В большинстве типов клеток морфологические изменения, связанные с апоптозом, в конце концов завершаются конденсацией хроматина, после чего следует распространенное расщепление ядра ДНК на олигонуклеосомные фрагменты. Кроме того, полученные ранее на линиях лимфоцитов и трансформированных клеток данные свидетельствуют, что внутриядерной фрагментации ДНК предшествует начальное расщепление хроматина на фрагменты с высокой молекулярной массой из 200-250 тыс. и/или 30-50 тыс. пар оснований.
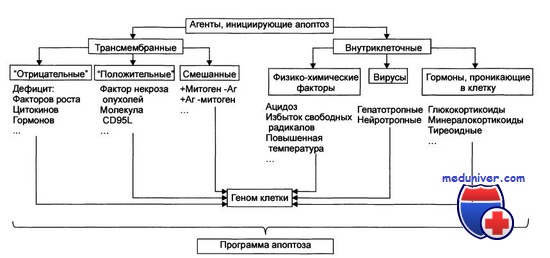
Стадия инициации апоптоза клеток
Вслед за этим после массивной эндонуклеотической деградации на олигонуклеосомные фрагменты идет дальнейшая деградация фрагментов из 50 тыс. пар оснований в характерную цепную структуру. Совокупные данные наводят на мысль, что латентные эндонуклеазы, способные к внутриядерному расщеплению, существуют в ядрах неапоптотических клеток и что именно депрессия нуклеазы наделяет различные агенты, индуцирующие апоптоз, клетка-специфической чувствительностью.
Представляет интерес для понимания ремоделирования хроматина и модификации гистонов тот факт, что эти процессы в дальнейшем регулируются NADPH-зависимыми гистонмодифицирующими ферментами. На лимфоцитарных клетках было показано, что воздействие покоящихся лимфоцитов на агенты, способные либо увеличивать скорость обрыва цепи ДНК, либо ингибировать разрыв и репарации ДНК, аналогичным образом вызывает ускорение потребления NAD.
Формирование внутриклеточных разрывов цепочки ДНК активирует механизмы репарации ДНК, которые связаны с поли-(АДФ)-рибозилированием нуклеарных белков с помощью ядерной поли-(АДФ-рибоза)-синтетазы. Нуклеарный фермент поли-(АДФ-рибоза)-синтетаза катализирует перенос АДФ-остатка NAD на существующие нуклеарные белки, что приводит к снижению аффинности связывания ДНК и последующему подавлению апоптоза.
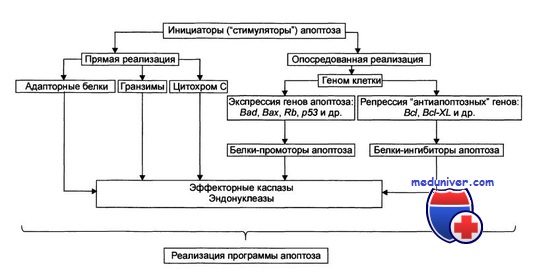
Стадия программирования апоптоза клеток
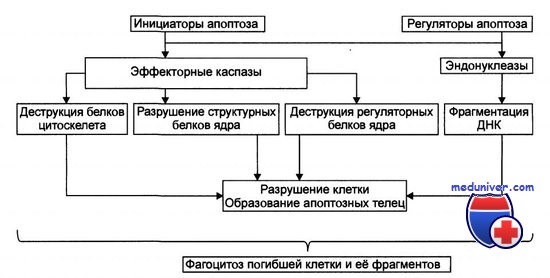
Сигнальная трансдукция и апоптоз клетки
В ответ на внутренние или внешние сигналы клетки пытаются восстановить свою структуру и организовать защиту. Если это им не удается, клетки гибнут вследствие апоптоза. В конечном счете каждый из этих путей подает сигналы посредством супрессора опухолей р53, который действует как ключевой интегратор апоптотических сигналов. Супрессор р53 рассматривается в качестве «охранителя генома».
В его функции входит распознавание и интегрирование идущих изнутри сигналов о повреждении клеток, чтобы включить либо остановку роста для «ремонта» поврежденной ДНК, либо апоптоз, чтобы индуцировать гибель клетки. В итоге эти сигналы должны сообща замыкаться на митохондриях, регулируя либо механизмы «ремонта» клетки, либо эндонуклеаза-зависимый процесс апоптоза, как это было описано ранее.
До того как все процессы сойдутся на митохондриях, важным шагом в ингибировании или индукции апоптоза является баланс про- и антиапоптотических механизмов. В настоящее время установлено, что семейство белков Вс1-2 в конечном счете управляет митохондриями с различным регулированием их проапоптотической ветви (порообразующих белков, например Вах, ВАК, BID, PUMA, NOXA и BIM) и антиапоптотической ветви (например, Вс1-2 и Bc1XL).
Координированным функционированием этих белков является сигнальная трансдукция на апоптоз активатора Мус. Достаточно сказать, что Мус может сенсибилизироваться к апоптозу в любой момент клеточного цикла в отсутствие трансляции de novo в результате своей способности функционировать в качестве активатора транскрипции апоптотических активаторов или их репрессоров.
Процессы, идущие вниз по ходу трансляции, запускают или сдерживают перемещение Вах с его эндогенной локализации в цитоплазме к внешней мембране митохондрий. После перемещения и гомоолигомеризации из митохондрий высвобождается цитохром-с, который затем функционирует через связывание Apaf-1 и опосредованный расщеплением каскад активации протеаз семейства каспаз. В итоге каспазы сходятся на критических клеточных субстратах, включающих поли-(АДФ-рибоза)-полимеразы, актин и pRb.