Саморегуляция чувствительности эффектора к гормональному сигналу. Десенситизация ( снижение чувствительности ) клетки.
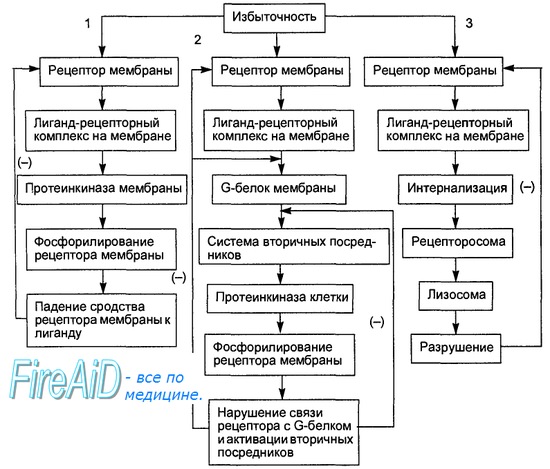
Изменение характера и интенсивности регуляторных сигналов, поступающих к клетке, ведет к адаптивной перестройке ее рецепторных структур за счет специальных механизмов саморегуляции, изменяющих чувствительность клетки к этому сигналу. Так, при избыточности внеклеточного гуморального сигнала формируется десенситизация соответствующего рецептора, т. е. уменьшение его чувствительности к гуморальному регулятору из-за ослабления сродства рецептора к молекуле лиганда или числа рецепторных молекул. Механизм десенситизации рецепторов, реализующих регулятор-ный эффект через системы вторичных посредников, представлен на рис. 6.4. При наличии во внеклеточной среде избыточного гуморального сигнала и/или увеличении времени связывания лиганда с рецептором активируемая лиганд-рецепторным взаимодействием протеинкиназа плазматической мембраны фосфорилирует рецептор, в результате его аффинность к лиганду уменьшается в 2—5 раз, что предотвращает чрезмерность регуля-торного эффекта. Если этого снижения чувствительности рецептора недостаточно, рецептор фосфорилируется другой протеинкиназой, активируемой соответствующим вторичным посредником. Следствием такого фос-форилирования является нарушение сопряжения рецепторов с G-белками и нарушение эффектов G-белков (активирующих или ингибирующих) на системы вторичных посредников и ионные каналы. Наконец, в случаях недостаточной эффективности двух указанных механизмов десенситизации гормон-рецепторные комплексы интернализуются внутрь клетки с образованием рецепторосомы (вакуоль с рецептором). В последующем, если уровень гуморального регулятора во внеклеточной микросреде снизится в течение 2—3 ч, интернализированный рецептор может вновь встроиться в мембрану, а если повышенная концентрация регулятора сохраняется — рецепторосомы сливаются с лизосомами и рецепторы разрушаются. Десенситизация рецепторов может быть гомологической, проявляющейся в снижении чувствительности только к тому гуморальному регулятору, избыток которого ее вызвал, и гетерологической, заключающейся в снижении чувствительности и к другим гуморальным факторам регуляции, но также реализующим эффект через связанные с G-белком рецепторы и те же вторичные посредники.
Чувствительность клеток к внешним регуляторным сигналам зависит также и от числа так называемых свободных, или резервных, рецепторов, т. е. доступных для лиганда. Увеличивая синтез этих мембранных структур или их «всплывание» на поверхность мембраны, клетка способна повышать чувствительность к регуляторному сигналу (сенситизация), например в случаях недостаточного количества молекул гуморального регулятора во внеклеточной микросреде. Повышение чувствительности к регуляторному сигналу является также следствием возросшего сопряжения рецепторов с G-белком и системой вторичных посредников.
Рис. 6.4. Схема трех путей десенситизации клетки к избыточному внешнему регуляторному сигналу. 1. Снижение сродства рецептора к лиганду из-за фосфорилирования рецептора мембранной протеинкиназой. 2. Нарушение связи молекулы рецептора с G-белком при фосфорилирова-нии рецептора цитоплазматическими протеинкиназами. 3. Интернализация лиганд-рецептор-ного комплекса и разрушение рецептора ферментами лизосом. Стрелками со знаком (—) обозначено подавление процесса взаимодействия.
Клетки организма, как правило, используют не один, а несколько путей реализации одного гуморального регуляторного сигнала, в связи с чем для каждого лиганда (например, молекул одного гормона) на мембранах существуют несколько типов рецепторов. Так, для ацетилхолина, гистамина, катехоламинов, опиоидных пептидов, серотонина, вазопрессина идентифицировано два (вазопрессин), три (опиоиды), четыре (мускариновые для ацетилхолина) и более (катехоламины, серотонин) мембранных рецепторов. Соответственно, меняя за счет саморегуляции чувствительность рецепторов одного типа, клетка изменяет характер функционирования и способна компенсировать неадекватные сдвиги в регуляторных влияниях.
Одним из регуляторных эффектов, реализуемых внутри клетки с помощью систем вторичных посредников мембранного лиганд-рецепторного взаимодействия, является изменение функционального состояния генома клетки и биосинтеза белка. Но и ядро клетки способно менять состояние плазматической мембраны, ее чувствительность к сдвигам внеклеточной среды, в том числе и изменению в ней количества и характера регуляторных сигналов. В клетках под контролем генома происходит синтез специальных белков — инверторов, оказывающих различное влияние на функциональное состояние мембраны в зависимости от адаптивно-компенсаторных потребностей клетки — изменение активности рецепторов, состояния ионных каналов и насосов. Эти белки могут вести к гиперполяризации клеточной мембраны, ослаблению функциональной активности клетки, с одновременным повышением синтеза белка и процессов регенерации внутриклеточных структур.