Влияние гипофиза на менструальный цикл. Гонадотропины - ФСГ и ЛГ
Гипофиз состоит из двух долей: передней (аденогипофиз) и задней (нейрогнпофиз). Так называемая промежуточная часть гипофиза у человека редуцирована и представляет узкую полоску, различимую только под микроскопом. Гипофиз расположен в турецком седле и ножкой соединен с гипоталамусом и остальными отделами центральной нервной системы; он составляет с гипоталамусом единый комплекс. Целесообразно различать [Баранов В. Г.], основываясь на функционально-анатомических особенностях, два звена в гипоталамо-гипофизарном комплексе: 1) гипоталамо-аденогипофизарную систему и 2) гипоталамо-нейрогипофизарную железу внутренней секреции. На уровне первого звена происходит секреция рилизинг-гормонов, вызывающих стимуляцию или торможение секреции гормонов аденогипофиза. Второе звено складывается из нейросекреторных ядер гипоталамуса, супраоптико-гипофизарного тракта и нейрогипофиза.
Нейрогипофиз не является железой внутренней секреции, так как биосинтез вазопрессина и окситоцина осуществляется в супраоптических и паравентрикулярных ядрах гипоталамуса; в нейропшофизе происходят только накопление этих гормонов и выделение их в кровь.
На менструальный цикл влияют гормоны передней доли гипофиза: тиротропин — тиротропный (ТТГ), соматотропный гормон (СТГ), кортикотропин — адренокортикотропный (АКТГ), лактотропин — лактотропный (ЛТГ), или пролактин, фоллитропин — фолликулостимулирующий (ФСГ), лютропин — лютеинизирующий (ЛГ), мелапотропин — меланостимулирующий (МСГ) и липотропный (ЛИГ) гормоны. Эти гормоны по химическому строению могут быть разделены на три группы: 1) простые белки (гормон роста, пролактин), 2) глюкопротеиды (фолликулостимулирующий, лютеинизирующий и тиреотропный гормоны) и 3) полипептиды (адренокортикотропный, меланостимулирующий и липотропный гормоны).
Дистальная часть передней доли гипофиза состоит из многочисленных эпителиальных перекладин (трабекул), в пространствах между которыми содержится большое количество капилляров, рыхлой соединительной и ретикулярной ткани. В трабекулах различают два вида железистых клеток-аденоцитов — хромофобные и хромофильные. Хромофобные клетки находятся в центре, их 50—60%. Цитоплазма их слабо окрашена и содержит небольшое количество органелл. Хромофильные клетки расположены по периферии трабекул и содержат большое количество секреторных гранул.
Хромофильные клетки разделяются на ацидо- и базофильные. Ацидофильные (эозинофильные) клетки имеют овальную форму, цитоплазма их содержит много секреторных гранул, окрашивается азаном в розовый цвет. Базофильныс клетки крупные, овальной или полигональной формы.
По вопросу о функциональных особенностях отдельных видов аденоцитов существуют две противоположные точки зрения. Одни авторы считают, что отдельные виды аденоцитов не обладают специфической способностью продуцировать определенные гормоны; их отличие определяется лишь различным функциональным состоянием аденоцитов. Другая группа авторов, наоборот, приписывает каждой форме аденоцитов специфическую гормональную функцию. Согласно взглядам этих авторов, ацидофильные клетки делятся на две подгруппы: 1) клетки, связанные с продукцией пролактина — ЛТГ (так называемые лактопротоциты). Они расположены в центре железы и содержат крупные (до 600 нм) секреторные гранулы; 2) клетки, связанные с продукцией соматотропного гормона — СТГ (так называемые соматотропоциты), расположенные вдоль сосудов, с секреторными гранулами величиной до 350 нм.
Базофильные клетки делятся на три подтипа: 1) клетки, связанные с образованием фоллитропина — ФСГ (так называемые фолликулостимулирующие гонадотропины); они небольшой величины, округлой формы, секреторные гранулы размером до 200 нм; 2) клетки, ответственные за продукцию лютропина — ЛГ (так называемые лютеинизирующие гонадотропоциты); они располагаются ближе к центру железы, более крупных размеров; их называют дельта-базофильными аденоцитами (дельта-клетки); 3) клетки, ответственные за продукцию тиротропного гормона — ТТГ (так называемые тиротропоциты). Это бета-базофильные аденоциты (бета-клетки). Они крупного размера, расположены в центре железы, окрашиваются альдегидфуксином. Секреторные гранулы их мелкие — до 150 нм. Отростчатые клетки хромофобного ряда (так называемые кортикотропоциты) продуцируют адренокортикотропный гормон — АКТГ. Секреторные гранулы их невелики—200 нм.
Существуют два типа секреции гонадотропинов: тонический и циклический. Тоническая секреция гормонов осуществляется непрерывно на сравнительно невысоком (базальном) уровне. Циклическая секреция гормонов происходит в определенной фазе менструального (полового) цикла, и ее уровень гораздо выше уровня тонической секреции.
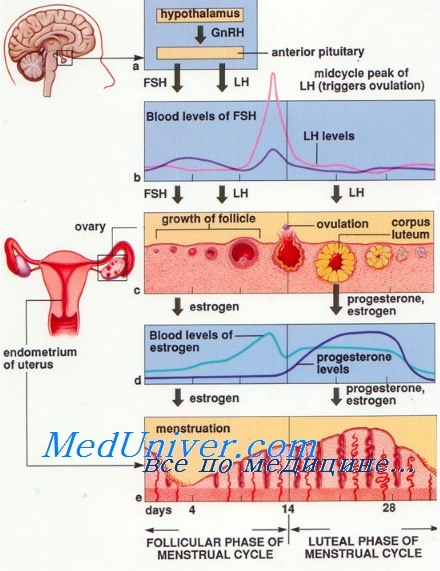
Данные о динамике содержания гормонов в суточной моче и крови приведены на рисунке. Аналогичные сведения об изменении содержания ЛГ и ФСГ в крови в различные фазы цикла получили Bart и Moreno. Так, уровень ЛГ в крови (при определении его методом радиоиммунодиффузии) колеблется незначительно в первой половине цикла, постепенно увеличиваясь ко времени овуляции. В ранней пролиферативной фазе уровень ЛГ в среднем составляет 10,6 мМЕ/мл, в поздней пролиферативной фазе—18,02 мМЕ/мл, в ранней секреторной фазе—10,5 мМЕ/мл. Уровень ФСГ в первой фазе выше, чем во второй; в первые дни после овуляции уровень ФСГ резко снижается.
Периодически наступающее повышенное выделение ЛГ в кровь связано с тем, что регуляция гонадотропных функций осуществляется двумя гипоталамическими образованиями [Алешин Б. В.]. Первое («низший центр»), представленное аркуатным и вентромедиальным ядрами, побуждает переднюю долю гипофиза к непрерывной тонической (базальной) секреции. Второе образование («высший центр»), располагающееся в преоптической области медиобазального гипоталамуса, оказывает модулирующее влияние на деятельность «низшего центра» и активизирует через него переднюю долю гипофиза к резкому повышению выделения ЛГ.
Регуляция менструального цикла обусловливается последовательным повышением секреции трех гонадотропных гормонов. Это способствует развитию и созреванию фолликула, овуляции и образованию желтого тела, сопровождается повышением секреции эстрогенов в первой фазе и повышением прогестерона во второй фазе менструального цикла. В первой половине цикла преобладает ФСГ, в середине цикла секретируются ФСГ и ЛГ, во второй половине его — ЛГ и ЛТГ.
Действие различных гонадотропинов на половую систему представляется довольно сложным. Так, оказалось, что ФСГ, вызывающий рост фолликулов в яичнике, в то же время только в сочетании с ЛГ может вызвать секрецию эстрогенов клетками растущего фолликула, лготсинизацию гранулезных клеток фолликула и увеличение размера матки [Lostroh, Johnson, Eshol]. В то же время только ЛГ не может вызвать роста фолликула, секреции эстрогенов гранулезными клетками, овуляции; при этом всегда необходимо наличие небольшого количества ФСГ (Israel). Специфическое действие ФСГ на яичники заключается в стимуляции митоза фолликулярных клеток и синтеза дезоксирибонуклеиновой кислоты (ДНК) в ядрах клеток (Callantine, Humphrey); для осуществления этого влияния необходимо одновременное действие эстрогенов.
ЛГ обеспечивает разрыв фолликула, выхождение яйцеклетки с последующим продвижением ее и имплантацией в эндометрий. ЛГ контролирует образование и функциональную активность желтого тела в яичнике человека. Самым начальным этапом действия ЛГ, вызывающего стимуляцию стсроидогенеза, является обратимое связывание его с белковыми рецепторами клеточной поверхности [Kammerman]. ЛГ связывается клеточной поверхностью, не проникая в клетку. Связываясь с рецептором, ЛГ активизирует аденилциклазу, находящуюся также в клеточной оболочке. В яичниках ЛГ связывается с интерстициальными клетками, клетками theca interna и гранулезы фолликула и желтого тела.
Учебное видео гормоны гипофиза в норме и при патологии
При проблемах с просмотром скачайте видео со страницы Здесь