Строма костного мозга. Роль стромы костного мозга в регуляции кроветворения.
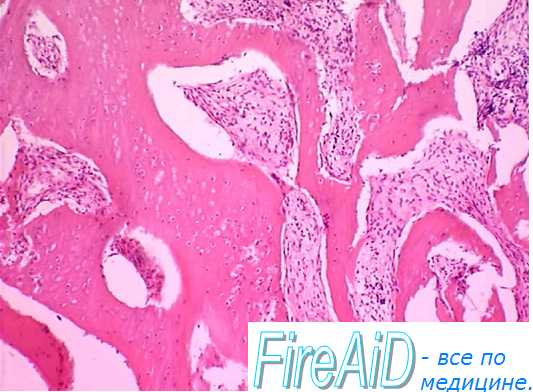
Строма костного мозга [макрофаги, соединительнотканная оболочка, выстилающая костномозговую полость (эндост), костномозговые синусоиды, жировые клетки, соединительная ткань и нервные окончания] создает «ге-мопоэтическое индуцирующее микроокружение», необходимое для пролиферации, дифференциации и фиксации СКК в костном мозге, размножения и созревания миелоидных клеток. Оно включает ростковые факторы, а также экстрацеллюлярный матрикс, содержащий фибронектин, ламинин, коллаген и гликозаминогликаны.
Делящиеся КОК взаимодействуют преимущественно с эндостом, являющимся источником ростовых факторов. Более зрелые клетки располагаются ближе к костномозговым синусоидам, что облегчает выход созревающих клеток крови через стенку синусоида в кровь.
СКК, КОК-ГММЭ и малодифференцированые бипотенциальные клетки постоянно выходят из костного мозга в кровь и транспортируются в другие участки кроветворной ткани. Это обеспечивает постоянный обмен клетками между анатомически разделенными участками кроветворной ткани. Однако пролиферировать и дифференцироваться в клетки крови кроветворные клетки-предшественницы способны лишь в ткани костного мозга, и необходимые для этого условия создает ГИМ. Так, адгезивный гликопротеин — фибронектин выполняет «якорную», фиксирующую функцию в отношении КОК-ГММЭ и эритроидных КОК и одновременно увеличивает их пролиферацию. Особые же микроворсинки ретикулярных и эндотелиальных клеток стромы фиксируют СКК и КОК-ГМ.
Строма костного мозга продуцирует КСФгм, КСФг, КСФмон, ИЛ-6, а также особый фактор — модулятор, «фактор стволовой клетки», который не вызывает пролиферации КОК, но резко усиливает пролиферативный эффект КСФ на эти клетки. При дефекте его воспроизводства стромой возникает нарушение кроветворения.
Нейтральные и кислые гликозаминогликаны увеличивают концентрацию цитокинов в непосредственной близости от клеток-мишеней, а также проницаемость мембран гемопоэтических клеток для кальция. Цитокины активируют в КОК формирование гемопоэтических вторичных мессенджеров, получивших название «сигнальные трансдукторы и активаторы транскрипции» (STAT), «ядерные факторы» (NF-E1, NF-E2, NF-kb/REL).
Вторичные мессенджеры обеспечивают быстрое распространение сигнала от возбужденного цитокином рецептора гемопоэтической клетки к ее геному, активируют транскрипцию его участков, ответственных за специализацию клетки-предшественницы, т. е. дифференциацию ее в направлении определенной клеточной линии — эритроидной, моноцитарной, нейтро-фильной и т. д. Нарушение формирования вторичных мессенджеров в КОК, не позволяющее цитокину реализовать регулирующий эффект на кроветворную клетку-предшественницу, приводит к нарушениям кроветворения. Так, нарушение, вызванное мутацией в участке генома, ответственного за синтез одного из транскрипционных факторов — NF-E1 обрывает развитие и влечет гибель эритроидных клеток на стадии проэритроб-ластов.