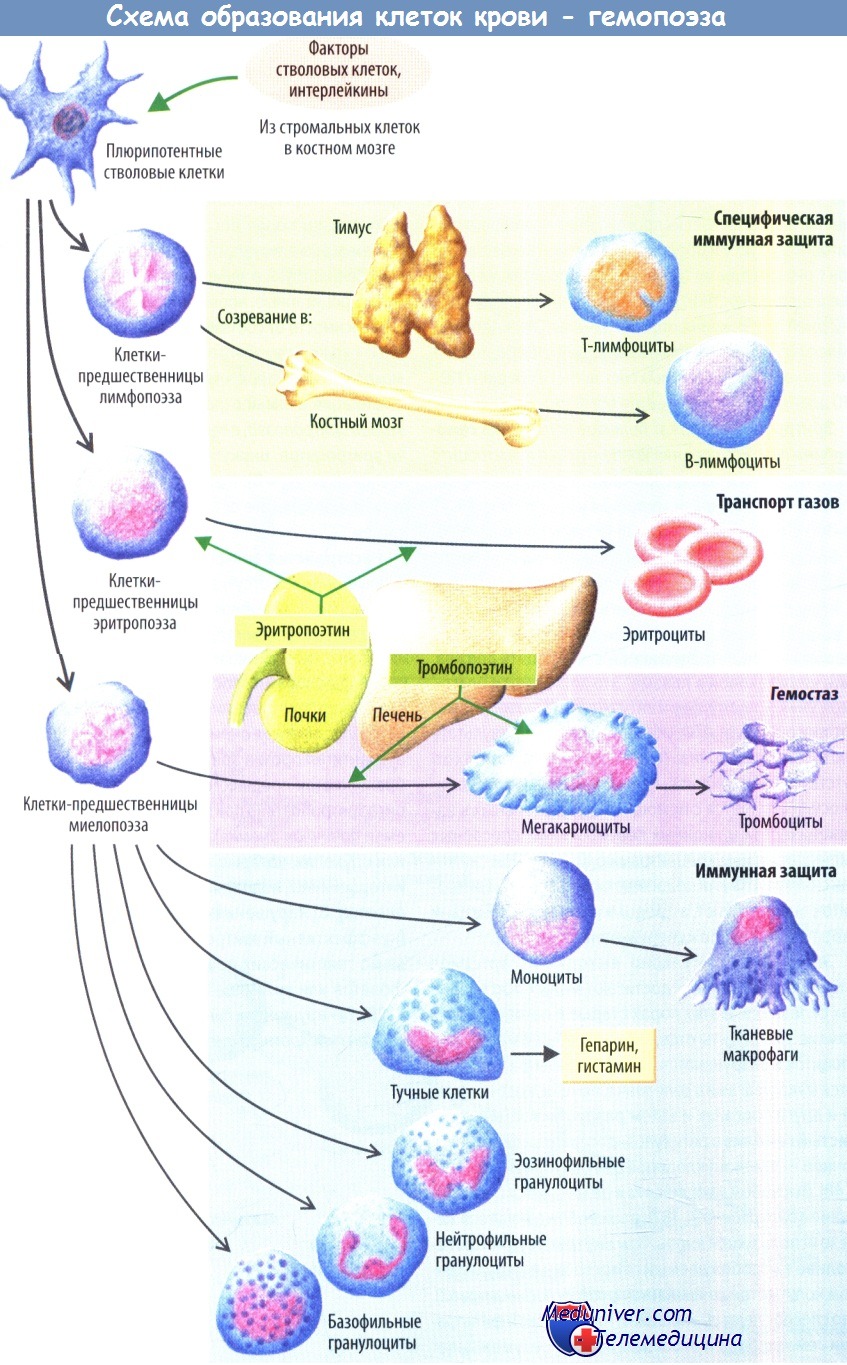
Рост различных КОК в культурах удается получить, добавляя стимулирующие его факторы. Отсюда факторы, стимулирующие образование грануло-цитарных колоний, получили название «колониестимулирующие факторы гранулоцитарные» — КСФ-Г, макрофагальные — КСФ-М, гранулоцитарно-макрофагальные — КСФ-ГМ, эозинофильные — КСФэоз (ИЛ-5), мегакариоцитарные — КСФмег (тромбопоэтин), стимулирующие развитие колоний из КОК-ГММЭ — мульти-КСФ или интерлейкин-3 (ИЛ-3).
ИЛ-3 и КСФ-ГМ стимулируют пролиферацию и дифференциацию не только КОК-ГММЭ, но и бипотентных КОК—КОК-ГМ, КОК-нейтрофильно-эозинофильных и др. В связи с этим ИЛ-3 и КСФ-ГМ рассматриваются как неспецифические факторы, поддерживающие самообновление и пролиферацию КОК-ГММЭ и бипотентных КОК. Все КСФ гликопро-теины относятся к полипептидным гормонам, которые регулируют гемо-поэз. Источниками ИЛ-3 и КСФэоз (ИЛ-5) являются Т-лимфоциты и спленоциты, КСФ-ГМ — Т-лимфоциты, моноциты, эндотелиальные клетки и фибробласты, КСФ-Г, КСФ-М-моноциты и фибробласты, а эритро-поэтина — перитубулярные и тубулярные клетки почек, купферовские клетки печени. Эти вещества получили общее название «гемопоэтические цитокины».
Гены, контролирующие синтез ИЛ-3, КСФ-Г и КСФ-М у человека, находятся на дистальной части длинного плеча 5-й хромосомы, КСФ-Г — локализованы у человека на 17-й хромосоме, эритропоэтины — на 7-й хромосоме.
Тяжелые нарушения в системе крови могут возникать при повреждении участков генома, ответственных за синтез молекулярных регуляторов гемопоэза, за синтез их рецепторов на КОК. Так, например, причиной тяжелой наследственной гранулоцитопении (нейтропении детей), приводящей их к ранней гибели из-за легкой поражаемости микробными и вирусными инфекциями, является нарушенное воспроизводство КСФ-Г в организме больных. Гранулоцитопоэз и уровень нейтрофилов в крови быстро восстанавливается при введении КСФ-Г этим больным. Точечная мутация над-мембранного района рецепторов эритропоэтина вызывает их резкую активизацию даже при малых количествах эритропоэтина в крови, что приводит к усиленной пролиферации эритроидной ткани и развитию эритроцитоза.
Продукцию КСФ усиливают различные стимулы: гипоксия — эритропоэтина; тромбоцитопения — тромбопоэтина; микробная инфекция — КСФ-ГМ, КСФ-Г, КСФ-М; гельминтная инфекция — КСФэоз (ИЛ-5). Каждый из перечисленных стимулов одновременно вызывает продукцию неспецифических ростковых факторов ИЛ-3 и КСФ-ГМ. При этом ИЛ-3 и КСФ-ГМ стимулируют как пролиферацию КОК-ГММЭ и КОК-бипотент-ных, так и формирование на их мембране рецепторов к КСФ, действие которых направлено на унипотентные КОК. Поэтому высокие концентрации КСФ-Г, КСФ-М и т. д. стимулируют пролиферацию и дальнейшую дифференциацию унипотентных клеток-предшественниц гранулоцитарного, моноцитарного и других рядов.
На интенсивность продукции КСФ оказывает регулирующее влияние вегетативная нервная система. Так, введение животным а- и b-адренобло-каторов значительно уменьшает уровень КСФ в крови. Стимуляторами ге-мопоэза являются Т-лимфоциты (Т-хелперы). Действие на организм возбуждающих гемопоэз факторов (кровопотеря, высотная гипоксия и др.) вызывает миграцию лимфоцитов в костный мозг и активацию ими КОК. КСФ регулируют функции не только КОК, но и зрелых клеток крови. Так, КСФ-ГМ усиливает фагоцитарную активность, метаболизм, миграцию в ткани зрелых нейтрофилов и моноцитов-макрофагов. КСФ-Г стимулирует бактерицидную, фагоцитарную и цитотоксическую активность этих клеток.
ИЛ-3 также усиливает цитотоксическую активность макрофагов, увеличивает эозинофильную фагоцитарную активность, а КСФ-М — бактерицидную и тумороцидную (разрушающую опухолевые клетки) функции моноцитов и макрофагов. Ответ клеточных линий кроветворной ткани на стимул внешней среды реализуется через рецепцию не только гемопоэти-ческих цитокинов, но и медиаторов нервных окончаний, находящихся в костномозговой ткани. Так, в паренхиме костного мозга представлены симпатические и пуринергические нервные окончания. Первые из них секретируют адреналин, норадреналин, дофамин, вторые — нейрокини-ны — субстанцию Р и др. Рецепторы этих соединений представлены на КОК-ГММЭ, КОК-ГМ, КОК-Г, КОК-ЭРМЕГ, КОК-Э.
Нейрокинины, связываясь с рецепторами на КОК, могут как усиливать, так и тормозить эффект гемопоэтических цитокинов на кроветворные клетки-предшественницы.
Например, связывание субстанции Р с рецепторами клеток-предшественниц активирует взаимодействие различных гемопоэтических цитокинов (ИЛ-3, ИЛ-6, КСФ-ГМ, ФСК и др.) с рецепторами этих цитокинов на указанных клетках-мишенях, вызывая сигнал трансдукции, направленный к геному клетки с последующей активацией пролиферации и дифференциации клеток. Напротив, нейрокинин А, секретируемый клетками стромы, связываясь с другими рецепторами этих же кроветворных клеток-предшественниц, взаимодействует с трансформирующим ростовым фактором b, снижая пролиферативный эффект кроветворных клеток. Это свидетельствует, что регуляция гемопоэза осуществляется в интересах целого организма и приспособительные реакции кроветворной ткани обеспечиваются не только цитокинами, но и передачей к ней нервными волокнами регулирующих сигналов.
В регуляцию гемопоэза «включен» механизм ингибирования гуморальной обратной связью клеток-предшественниц продуктами, образуемыми клетками разных линий на последних этапах их созревания. Кислый изо-ферритин, продуцируемый зрелыми нейтрофилами, тормозит их новообразование из КОК-Г; образуемый альфа-гранулами тромбоцитов «трансформирующий рост фактор-b» угнетает развитие КОКмег; простагландины серии Е, а- и b-интерфероны, продуцируемые макрофагами, тормозят пролиферацию КОКмон; вещества ядер, вытолкнутых нормобластами, кейло-ны, образуемые зрелыми эритроцитами, тормозят дифференциацию КОК-Э. К цитокинам, тормозящим гемопоэз, относят опухольнекротизирующий фактор (ОНФ). Его рецепторы представлены на мембране различных КОК. Активация рецепторов ОНФ, в частности fas-рецептора опухольнек-ротизирующего фактора, на КОК вызывает выраженное угнетение гемопоэза и «запускает» апоптоз-запрограммированную смерть клетки-предшественницы. Fas-опосредованный апоптоз стволовых кроветворных клеток лежит в основе ряда наследуемых форм недостаточности костного мозга.