Фосфолипиды как вторичные посредники. Система кальций-кальмодулин как вторичный посредник
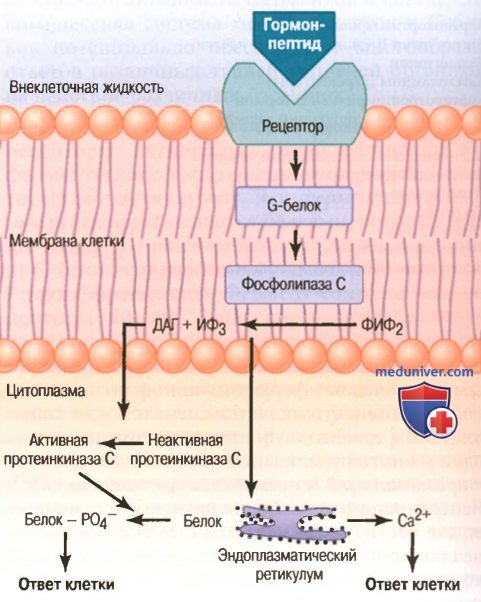
а) Фосфолипиды клеточных мембран как система вторичного посредника. Некоторые гормоны взаимодействуют с трансмембранными рецепторами, активирующими фермент фосфолипазу С, прикрепленную к внутренней поверхности рецепторов (для облегчения понимания просим вас изучить таблицу ниже).
Этот фермент катализирует распад некоторых фосфолипидов в мембране, особенно фосфатидилинозитолбифосфата, на два вторичных посредника: инозитолтрифосфат и диацилглицерол. ИФз мобилизует ионы кальция из митохондрий и эндоплазматического ретикулума, затем ионы кальция ведут себя как вторичные посредники, вызывая сокращения гладких мышц и секрецию в клетке.
Диацилглицерол (другой липидный вторичный посредник) активирует фермент протеинкиназу С, которая затем фосфорилирует большое количество белка, формирующего ответ клетки (для облегчения понимания просим вас изучить рисунок ниже).
Фосфолипиды мембран как система вторичных посредников, обусловливающая реализацию гормонального управления функциями клетки. ДАГ - диацилглицерол, ИФ3 - инозитолтрифосфат, ФИФ2 - фосфатидилинозитолбифосфат
Дополняет этот эффект липидная часть ДАГ — арахидоновая кислота, являющаяся предшественником простагландинов и других местных гормонов, обусловливающих многочисленные реакции тканей.
б) Система кальций-кальмодулин как вторичный посредник. Другая система вторичного посредника запускается вхождением кальция в клетку, которое может быть обусловлено:
(2) взаимодействием гормона с рецептором, открывающим кальциевые каналы.
Попав в клетку, ион кальция связывается с белком кальмодулином. Этот белок имеет четыре участка взаимодействия с кальцием. Когда три или четыре участка оказываются связанными с кальцием, кальмодулин меняет форму, инициируя активацию или ингибирование протеинкиназ наряду с прочими многочисленными эффектами. Активация кальмодулин-зависимой протеинкиназы совместно с фосфорилированием является причиной активации или ингибирования белков, вовлекаемых в ответ клетки на действие гормона. Например, одной из специфических функций кальмодулина является активация миозинкиназы, которая действует непосредственно на миозин гладких мышц, вызывая мышечные сокращения.
Нормальная концентрация кальция в большинстве клеток организма составляет 10-8-10-7 моль/л, что недостаточно для активации кальмодулиновой системы. Как только концентрация ионов достигает 10-6—10-5 моль/л, она становится достаточной для обнаружения всех проявлений активности кальмодулина. Это соответствует количественным изменениям концентрации ионов кальция, необходимых скелетной мышце для активации тропо-нина С, который обусловливает сокращение скелетной мышцы. Интересно, что тропонин С подобен кальмодулину как функционально, так и структурно.
в) Гормоны, действующие главным образом на генетический аппарат клетки. Стероидные гормоны увеличивают синтез белков. Другим способом действия гормонов, особенно стероидных, секретируемых корой надпочечников, яичниками и семенниками, является изменение синтеза белков в клетках-мишенях. Затем эти белки функционируют как ферменты, транспортные или структурные белки, что в целом обеспечивает изменение функций клеток.
1. Стероидный гормон диффундирует через мембрану клетки и попадает в цитоплазму, где связывается с белком-рецептором.
2. Комплекс гормона с белком-рецептором транспортируется или диффундирует в ядро.
3. Комплекс присоединяется к специфическим участкам ДНК хромосомы, активируя процессы транскрипции путем образования мРНК.
4. мРНК диффундирует в цитоплазму, где обеспечивает процессы трансляции на рибосомах, формирующих новые белки.
Например, альдостерон (один из гормонов, секретируемых корой надпочечников) попадает в цитоплазму клеток почечных канальцев, содержащих специфический белок-рецептор альдостерона.
Таким образом, в этих клетках мы наблюдаем уже изложенную последовательность событий. Приблизительно через 45 мин в клетках тубулярного аппарата начинают появляться белки, обеспечивающие реабсорбцию натрия и секрецию калия в канальцах.
Максимальная активность стероидных гормонов задерживается на срок от 45 мин до нескольких часов и даже дней. Это заметно отличается от почти немедленного действия некоторых гормонов-пептидов или производных аминокислот, таких как вазопрессин или норадреналин.