Сенсорные зоны коры головного мозга: соматосенсорная кора, теменная долька, зрительная кора, слуховая кора
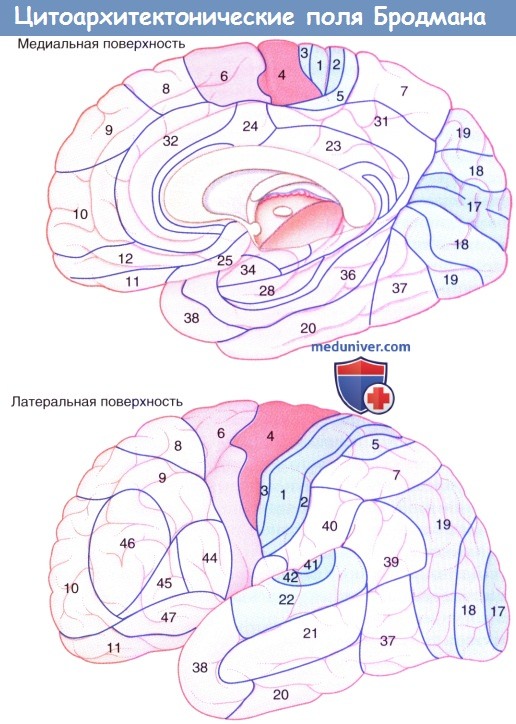
Цитоархитектонические поля Бродмана. Окрашенные зоны:
а) Моторная (красная):
4 — первичная моторная кора
6 — на медиальной поверхности, дополнительная моторная зона
6 — на латеральной поверхности, премоторная кора
б) Сенсорная (голубая):
3/1/2 — первичная соматосенсорная кора
40 — вторичная соматосенсорная кора
17 — первичная зрительная кора 18,
19 — ассоциативная зрительная кора
41, 42 — первичная слуховая кора*
22 — ассоциативная слуховая кора
(*Первичную слуховую кору не всегда можно увидеть сбоку, так как она полностью расположена на верхней поверхности верхней височной извилины.)
а) Соматосенсорная кора:
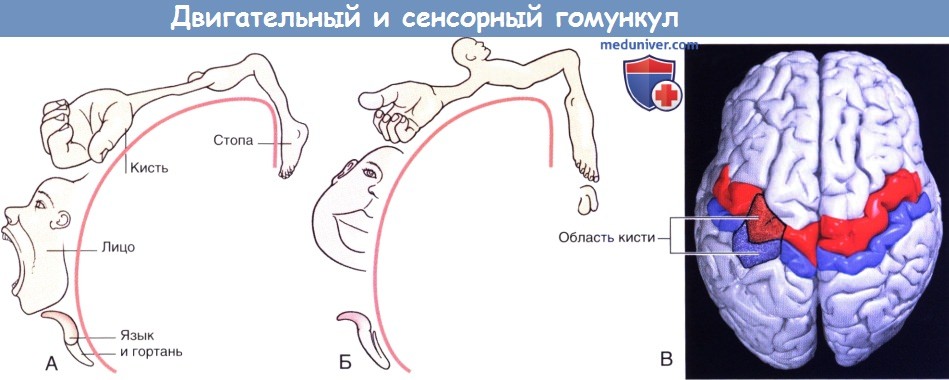
1. Отделы. Соматосенсорная, или соматоэстетическая, кора занимает всю постцентральную извилину. Части тела противоположной стороны представлены инвертированными (за исключением лица), а кисти рук, губы и язык представлены непропорционально большими. Представленные на рисунке ниже уже известные Вам гомункулы лишь схематически изображают расположение различных частей тела без учета обширных участков, перекрывающих друг друга.
На вертикальном срезе соматосенсорная кора разделена на поля 3, 1 и 2. Таламо-корковые волокна (от вентральных задних медиального и латерального ядер) подходят преимущественно к полю 3 (разделенному на более мелкое поле За и более крупное поле 3б) и в меньшей степени — к полям 1 и 2. Происходит распределение информация от кожных рецепторов, и быстро проводящие волокна направляются преимущественно к полю 1; к более сложно организованному полю 2 приходят импульсы от кожных и других рецепторов. Размер и сложность организации рецепторного поля прогрессивно увеличиваются от 3б до 1. Поля 3 (За иногда включают в состав моторной коры), 1 и 2 рассматривают как первичную соматосенсорную кору (SI), однако поле 3б больше «заслуживает» определение первичный. Нейроны сенсорной коры обладают постоянно изменяющимися функциональными свойствами и не просто производят извлечение соответствующей сенсорной информации, а систематизируют ее в зависимости от контекста или ситуации. Подобная контекстуальная обработка обусловливает адаптирующееся целенаправленное поведение; многократная обработка — основа обучения.
(А) Рисунок (адаптирован из Penfield и Rasmussen), изображающий перевернутого «двигательного» гомункула в левой прецентральной извилине, за исключением лица. Зоны, перекрывающие друг друга, не показаны.
(Б) Рисунок (адаптирован из Penfield и Rasmussen), изображающий перевернутого «сенсорного» гомункула в левой постцентральной извилине, за исключением лица. Зоны, перекрывающие друг друга, не показаны.
(В) Первичная двигательная кора (красная) и первичная соматосенсорная кора (синяя), вид сверху. Относительно более крупные моторные и сенсорные области левого полушария характерны для правшей.
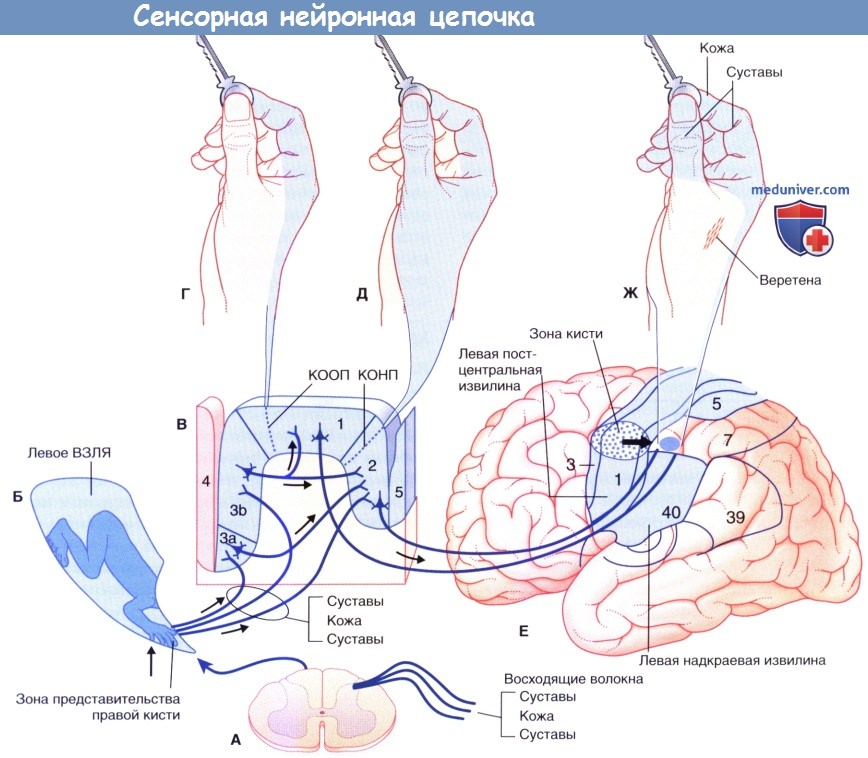
Сенсорная нейронная цепочка, позволяющая идентифицировать ключ только с помощью ощупывания.
(А) Закодированная сенсорная информация от правой кисти входит в спинной мозг и затем направляется по чувствительным нейронам второго порядка (противоположной стороны)
в медиальном отделе левого заднего канатика в составе медиального лемнискового пути в верхний отдел ствола мозга.
(Б) Зона, отвечающая за кисть в вентральном заднем латеральном ядре таламуса (ВЗЛЯ), содержит тела третичных чувствительных нейронов.
(В) Нейроны третьего порядка посылают волокна к зонам 3, 1 (опосредованно) и 2 соматосенсорной коры.
(Г) Корковый отдел «одного» пальца (КООП).
(Д) Корковый отдел нескольких пальцев (КОНП).
(Е) Левая теменная доля (обведенная область обозначает зону кисти), вид снаружи.
К зоне 7 подходят короткие ассоциативные волокна от зон 1, 2 и 5; в ней происходит объединение информации от кожи, мышечных веретен и суставных капсул.
Взаимосвязи с хранилищем тактильной информации здесь и в зоне 5 позволяют создать образ ключа без помощи зрения.
2. Афферентные волокна. Помимо таламических афферентных волокон от вентрального заднего ядра, к соматосенсорной коре подходят также комиссуральные волокна от противоположной соматосенсорной коры через мозолистое тело и короткие ассоциативные волокна от прилежащей первичной моторной коры. Множество волокон от моторной коры служат коллатералями корково-спинномозговых волокон, идущих к переднему рогу спинного мозга, и участвуют в создании чувства веса при подъеме объектов (потерю этого чувства называют барагнозией).
Поражение соматосенсорной коры часто возникает при окклюзии ветви средней мозговой артерии, кровоснабжающей сенсорную кору. Корковый тип потери чувствительности в этом случае проявляется ослаблением чувствительности на противоположной стороне тела, особенно на предплечье и кисти (проявляющимся повышением порога возбудимости, ослаблением дискриминационного чувства, нарушением вибрационного чувства и чувства положения), а также нарушением распознавания более сложных видов чувствительности, несмотря на сохранность болевой, температурной и даже вибрационной чувствительности. Поражение может проявляться невозможностью распознавания знакомых объектов при помещении их в руку (астереогноз), невозможностью распознавать фигуры и цифры, рисуемые на руке (аграфестезия), или неспособностью опознать одновременно два тактильных стимула, приложенных к противоположным частям тела (экстинкция). Потерю способности распознавать размер и форму объектов в результате поражения на уровне между чувствительными рецепторами и корой обозначают термином стереоанестезия.
Сложные неврологические нарушения при повреждении теменной доли, обычно недоминантного полушария (чаще правого), обозначают общим термином агнозии (тактильная агнозия — неспособность распознать форму объекта при его ощупывании, анозогнозия—отрицание болезни или неврологических нарушений, аутопагнозия—неспособность к определению, ориентированию и распознаванию собственных частей тела). Повреждение теменной доли (чаще левой) может, кроме того, приводить к апраксии—невозможности совершения целенаправленных движений или правильного использования объекта, несмотря на сохранную моторику и осознанное восприятие.
3. Эфферентные волокна. К эфферентным волокнам соматосенсорной коры относят ассоциативные, комиссуральные и проекционные волокна. Ассоциативные волокна направляются к моторной коре на той же стороне к полю 5 и полю 40 (надкраевой извилине). Комиссуральные волокна проходят к соматосенсорной коре противоположных полушарий. Проекционные волокна опускаются в заднем отделе пирамидного пути и оканчиваются на вставочных нейронах сенсорных релейных ядер (вентральное заднее ядро таламуса той же стороны), а также проходят в заднем канатике к заднему рогу спинного мозга на противоположной стороне. Передача чувствительной информации по спинно-таламическому проводящему пути может ингибироваться (тормозными вставочными нейронами) при интенсивных физических нагрузках, тогда как проведение через задний канатик (в составе медиального лемнискового пути) может усиливаться (возбуждающими вставочными нейронами) при исследовательской активности, например при пальпации рельефной поверхности.
б) Ассоциативная соматосенсорная кора (поле 5). Этим термином обозначают поле 5, расположенное сразу за соматосенсорной корой. Активация большей части этой области происходит при совершении хватательных движений противоположной кистью под контролем зрения (дорсальный зрительный путь описан ниже).
в) Верхняя теменная долька (поле 7). Верхняя теменная долька функционально соответствует полю 7. Нижний отдел поля 7 получает информацию от полей 1, 2 и 5. После получения тактильной и проприоцептивной информации от кожи, мышц и суставов поле 7 активирует собственную «память», осуществляющую распознавание объектов, взятых в (противоположную) руку, позволяя, таким образом, идентифицировать объект без помощи зрения.
Верхнюю часть поля 7 составляют клетки зрительного проводящего пути, отвечающего на вопрос «Где?».
г) Нижняя теменная долька (поля 39 и 40). Нижняя теменная долька образована полями 39 (угловая извилина) и 40 (надкраевая извилина). Оба поля отвечают за речь — функцию преимущественно левого полушария, описанную в отдельной статье на сайте; нарушение речи, вызванное поражением мозга, носит название афазия. (Поражение правого полушария может приводить к развитию неспособности понимания или использования эмоций при устной речи — апрозодии.)
д) Межтеменная кора. Повышенная активация коры в стенках межтеменной борозды возникает при выполнении задач, требующих зрительно-двигательной координации (например, при приближении и захвате объектов, идентифицированных полем зрения противоположной стороны, с последующим зрительным и тактильным пространственным анализом). К этой зоне относят также теменное глазодвигательное поле.
е) Вторичная соматосенсорная кора. На медиальной поверхности теменной покрышки островка расположена небольшая вторичная соматосенсорная кора (SII). Она получает ноцицептивные пути от таламуса и ярко светится на ПЭТ-снимках головного мозга при периферической болевой стимуляции. SII, вероятно, действует совместно с SI при обработке дискриминационной чувствительности или локализации болезненного участка.
Пластичность соматосенсорной коры. Можно очень точно выделить зоны отдельных пальцев кисти в соматосенсорной коре обезьян посредством записи электрических ответов колонн кортикальных клеток на тактильную стимуляцию поочередно каждого пальца. Эти пальцевые карты могут быть другими при оценке периферической чувствительности, на что указывают следующие данные.
• Срединный нерв иннервирует ладонную поверхность латеральных трех с половиной пальцев кисти, а лучевой нерв иннервирует их тыльные поверхности. При повреждении срединного нерва увеличивается зона представления в коре тыльной поверхности кисти, захватывая зону ладонной чувствительности. Расширение зоны начинается через несколько часов и прогрессивно увеличивается в течение нескольких недель. При восстановлении функции срединного нерва, кортикальная карта возвращается к норме.
• При денервации среднего пальца соответствующая корковая зона в течение нескольких часов не отвечает на стимуляцию, а затем прогрессивно (в течение нескольких недель) замещается зонами II и IV пальцев.
• При постоянной стимуляции кожи подушечки пальца (например, при соприкосновении с вращающимся диском для приготовления гранул пищи) представление этого пальца в коре может увеличиваться в два раза в течение нескольких недель, возвращаясь к норме после прекращения эксперимента.
Эти исследования показывают, что соматосенсорные карты обладают пластичностью и изменяются при нарушении периферической иннервации. Чисто анатомический подход (например, разрастание нервных волокон в центральной или периферической нервной системе) не позволяет объяснить ранние изменения, развивающиеся в течение нескольких часов: наоборот, их следует рассматривать как основу сенсорной конкуренции.
ж) Сенсорная конкуренция. В сенсорных картах, созданных на уровне задних рогов, задних канатиков, таламуса и соматосенсорной коры, выявлены признаки наложения (анатомического перекрытия). Например, таламо-корковые соматосенсорные зоны III пальца накладываются на зоны II и IV пальцев. В зоне наложения корковые колонны получают информацию от двух соседних пальцев. Вставочные корковые нейроны могут тормозить слабо возбужденные периферические колонны. В эксперименте (например, на кошках) число колонн, отвечающих на направленную таламо-корковую стимуляцию, может быть увеличено местным воздействием антагонистов ГАМК (бикукуллин), подавляющих периферическое торможение. Результат отсутствия периферического сенсорного поля может быть различным.
Если одна группа таламо-корковых нейронов перестает возбуждаться в результате потери сенсорной стимуляции, она больше не подвергается периферическому торможению, и корковые колонны на ее территории «захватываются» соседними, активными группами нейронов. В период расширения синаптические связи между клетками подвергаются как кратковременным, так и долговременным изменениям, что отражает процесс обучения.
На соматосенсорной карте тела человека пальцы расположены рядом с лицом. В нескольких документально подтвержденных случаях ампутации верхней конечности у пациентов впоследствии возникало ощущение «фантомного пальца» при прикосновении предметом к лицу на стороне ампутации (например, гребешком, зажатым другой рукой). Это ложное восприятие может появляться в течение двух недель после ампутации. Его можно объяснить прекращением предсуществующего наложения таламо-корковых нейронов.
з) Зрительная кора (поля 17, 18, 19). К зрительной коре относят первичную зрительную кору (поле 17) и ассоциативную зрительную кору.
1. Первичная зрительная кора (поле 17). Первичная зрительная кора — место окончания коленчато-шпорного тракта, через который передается информация от ипсилатеральных половин обеих сетчаток и, следовательно, от зрительного поля противоположного глаза. Этот миелинизированный путь образует в первичной зрительной коре бледные зрительные полоски (линии Геннари) перед созданием контактов с шиповатыми зернистыми клетками в высокозернистом слое IV. Зрительные полоски (впервые обнаруженные студентом-медиком Франческо Дженнари в 1775 г.) дали альтернативное название для поля 17 — полосатая кора.
Шиповатые зернистые клетки находятся в глазных доминантных колоннах, названных так, поскольку к чередующимся колоннам попеременно подходят импульсы от левого и правого глаз. Если можно было бы отдельно промаркировать волокна от каждого глаза и посмотреть на зрительную кору снаружи, эти чередующиеся колонны образовывали бы пучки в форме витков (наподобие отпечатков пальцев), где каждый из пучков отвечал бы на стимуляцию только одного из глаз. Коленчато-шпорный тракт представлен в коре таким образом, что соответствующие точки от двух сетчаток расположены в соседних колоннах на одной линии. Такое расположение идеально подходит для бинокулярного зрения, так как серия этих колонн образует модули, в краях которых происходит обработка информации от обоих глаз.
Недифференцированные импульсы от латерального коленчатого ядра «преобразуются» в ряд свойств в слое VI первичной зрительной коры. Этот процесс происходит благодаря распределению нейронов слоя VI в функциональные колонны. Взаимосвязи между этими нейронами, детально показанные на рисунке ниже, обеспечивают определение контуров предметов, их размер, направление движения и направленность зрительных раздражителей. Комплексная обработка происходит путем дальнейших корковых взаимодействий.
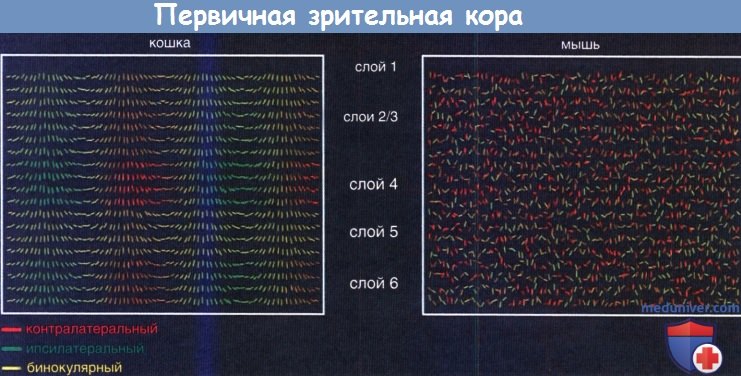
Представление избирательного действия нейронов первичной зрительной коры (V1), получающей информацию от латерального коленчатого тела.
Запись нейрональной активности взрослой кошки (слева) показывает высокую избирательность для специфических ориентаций стимулов (показаны наклоном линий) и различную степень доминирования противоположного (красный) и ипсилатерального (зеленый) глаз с множеством клеток, регулирующих деятельность обоих глаз.
Как ориентация, так и глазодоминирование (нейроны, в большей степени отвечающие на стимуляцию от одного глаза, чем от другого) распределены по колоннам. В колоннах, реагирующих на определенную ориентацию стимула, участвуют все клеточные слои, тогда как глазодоминирование происходит преимущественно в слое 4, где большое число клеток отвечает за монокулярное зрение. Поле V1 у мыши (справа) не имеет колончатого строения для ориентации или глазодоминирования.
Однако все же имеется высокая избирательность нейронов к ориентации, а также в некоторой степени отмечено глазодоминирование, но со смещением к противоположному глазу.
2. Пластичность первичной зрительной коры. Базовая схема и соотношение глазных доминантных колонок закладывается и устанавливается еще до рождения и сохраняется у животных, обитающих в полной темноте. При развитии в детстве слепоты на один глаз соответствующие корковые колонны остаются небольшими, а размеры корковых колонн видящего глаза увеличиваются.
3. Ассоциативная зрительная кора (поля 18 и 19). Ассоциативная зрительная кора представлена зонами 18 и 19, имеющими также общее название перистриарная, или экстрастриарная, кора. К ней подходят афферентные волокна преимущественно от поля 17, а также небольшое количество прямых таламических волокон от подушки таламуса. Клеточные колонны отвечают за обработку отдельных признаков. Часть колонн обрабатывает геометрические формы, часть отвечает за обработку цвета, другая часть — за бинокулярное (пространственное) зрение, а некоторые колонны — за более сложные задачи (распознавание лиц).
Большое количество перистриальных колонн имеют крупные рецепторные поля. Некоторые из них захватывают физиологическое слепое пятно (головку зрительного нерва) и могут отвечать за «маскирование» слепого пятна при монокулярном зрении.
Волокна, идущие от подушки таламуса к ассоциативной зрительной коре, считают частью проводящего пути, принимающего участие в «слепозрении» (остаточная обработка зрительной информации после разрушения первичной зрительной коры). Это удивительное состояние наблюдают у пациентов после тромбоза шпорной ветви задней мозговой артерии. Несмотря на полное отсутствие противоположного зрительного поля, эти пациенты, тем не менее, способны следить за движущимся пучком света без его восприятия, только лишь по «ощущению» его присутствия. Истинный проводящий путь остается неустановленным; к вероятным путям относят прохождение зрительных волокон через медиальный корешок зрительного тракта или верхний холмик, или от подушки таламуса к ассоциативной зрительной коре, или корковый путь от латерального коленчатого тела.
Наиболее функционально сложные модули зрительной ассоциативной коры расположены в латеральном и медиальном отделах поля 19. Латеральную группу модулей неформально относят к дорсальному «Где?» зрительному проводящему пути. Медиальная группа принадлежит к вентральному проводящему пути, отвечающему на вопрос «Что?»; оба проводящих пути функционируют одновременно, их не следует рассматривать отдельно друг от друга.
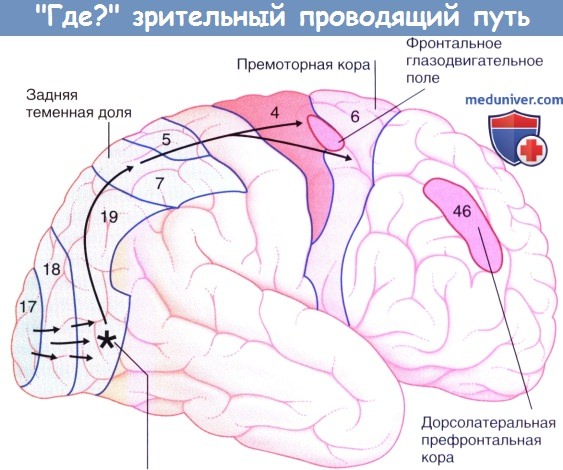
4. «Где?» зрительный проводящий путь. На основании записей электрической активности у испуганных обезьян и при ПЭТ-исследовании у людей-добровольцев было установлено, что латеральная часть поля 19 отвечает преимущественно за регистрацию движений, происходящих в противоположной половине поля зрения. Основная часть волокон от этого поля направляется к полю 7, обозначаемому клиницистами как задняя теменная кора. Помимо регистрации движений, поле 7 отвечает также за стереоскопическое (пространственное) зрение, которое вместе с пространственным чувством позволяет определять положение объектов относительно друг друга.
Поле 7 получает волокна от подушки таламуса, участвующие в «слепозрении», и взаимодействует через верхний продольный пучок с ипсилатеральным фронтальным глазодвигательным полем и премоторной корой.
Активация клеточных колонн поля 7 у обезьян происходит, когда важный объект (например, фрукт) появляется в противоположной половине зрительного поля. Через ассоциативные волокна возбужденные клеточные колонны повышают частоту фоновой активности колонн во фронтальном глазодвигательном поле и премоторной коре, но без инициации движения. Этот эффект называют скрытым вниманием, или скрытой ориентацией. Внимание (ориентация) становится открытым, когда животное отвечает саккадическими (высокочастотными содружественными движениями глаз) и хватательным движением в направлении объекта (или без него). При поражении поля 7 двигательные реакции на важные объекты возникают с опозданием, и хватательные движения противоположной руки становятся неточными.
При ПЭТ-исследовании у людей-добровольцев было установлено повышение коркового метаболизма в поле 7 в ответ на движение объекта в противоположной половине зрительного поля. При движении противоположной руки к объекту одновременно возбуждаются поля 5 и 7. У людей (как и у обезьян) поражение поля 7 сопровождается неуклюжими, неточными движениями в противоположном зрительном поле. Комплекс «Где?» является также комплексом «Как?», поскольку пространственно-зрительная информация используется двигательной системой для создания направленных движений.
При появлении в поле зрения объекта, представляющего особый интерес, у добровольцев активируются все дополнительные зоны коры. Дорсолатеральная префронтальная кора (ДЛПФК, ориентировочно соответствует полю 46 по Бродману) — важная зона принятия решений, особенно решения о приближении или отступлении, показана на рисунке ниже и также упомянута ниже. На отдельном рисунке показан участок коры передней поясной извилины. Эта область подробно рассмотрена в отдельной статье на сайте, но упомянута здесь в связи с тем, что ее возбуждение дорсолатеральной корой возникает, когда человек уделяет внимание зрительной задаче.
Латеральная поверхность правого полушария, показан ход зрительного «Где?» проводящего пути от зрительной коры к теменной и лобной долям.
Звездочкой указана область, отвечающая за обнаружение движений в левом зрительном поле.
Активность правого фронтального глазодвигательного поля облегчает саккадические движения к левому зрительному полю.
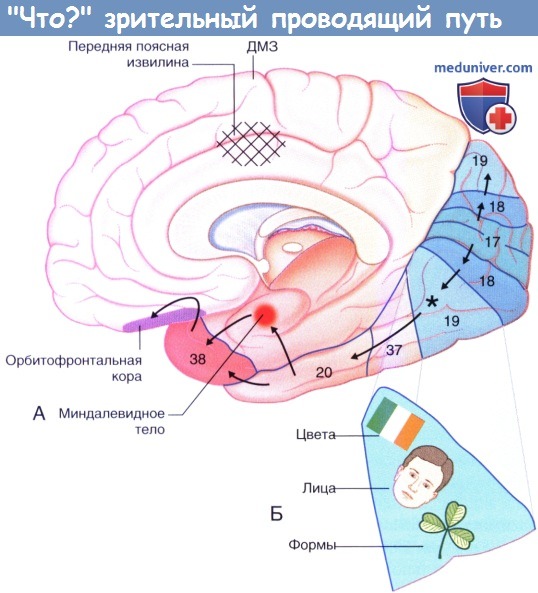
5. «Что?» зрительный проводящий путь. Вентральный зрительный проводящий путь соединяется с переднемедиальным отделом поля 19, расположенным преимущественно в веретеновидной извилине — части затылочно-височной извилины. Считают, что эта область участвует в распознавании трех типов зрительных признаков (нейроны этой зоны в дальнейшем обрабатывают данные признаки в зависимости от когнитивных и поведенческих аспектов), указанных на рисунке ниже.
• В относительно латеральном отделе расположены модули, определяющие форму всех типов объектов, включая форму букв. Это центр общей (категориальной/классической) идентификации объектов (например, собаки как таковой, без уточнения).
• В среднем отделе расположены модули, специфическая функция которых заключается в распознавании человеческих лиц.
• В относительно медиальном отделе находится отдел распознавания цвета, необходимый для определения всех цветов, кроме черного и белого. Ахроматопсия (цветовая слепота может развиться при поражении любого отдела зрительного проводящего пути) может возникать вследствие стойкого снижения артериального давления в обеих задних мозговых артериях, вызванного, например, эмболией в верхушку основного ствола базилярной артерии, сопровождающейся развитием инфаркта мозга. Такие пациенты видят мир черно-белым (в оттенках серого).
Распознавание отдельных объектов и лиц — функция переднего отдела «Что?» зрительного проводящего пути в нижней височной извилине (поле 20) и коре полюса височной доли (поле 38). Обе эти области активируются, например, при распознавании лица Марии или моей собаки. Неспособность к распознаванию лиц (тип агнозии, называемый прозопагнозией) считают частым и тревожным признаком болезни Альцгеймера, когда пациент прекращает узнавать лица членов семьи, несмотря на сохранение способности к распознаванию общих объектов.
Угрожающие взгляды или лица приводят к возбуждению зонами 20 и 38 миндалевидного тела, особенно в правом полушарии; правое миндалевидное тело, в свою очередь, возбуждает отвечающую за страх правую орбитофронтальную кору, выделенную сиреневым цветом на рисунке ниже.
Как происходит активация зрительной ассоциативной коры, например, при принятии решения найти яблоко в корзине с фруктами или определенное слово на странице текста? При ПЭТ-исследовании активация лобной доли происходила во всех случаях, когда для решения задачи требовались руки. При решении зрительной задачи, связанной с обработкой цвета и формы, особенно активна ДЛПФК. При зрительном поиске роль лобной коры сводится к активации зрительной памяти в ассоциативной зрительной коре и высвобождению соответствующих образов в момент поиска. Происходит также возбуждение переднего отдела поясной коры. Помимо поступления информации от первичной к ассоциативной зрительной коре по дорсальному и вентральному проводящим путям, имеется также «нисходящий» путь, опосредующий осознанное восприятие и поведенческие реакции (например, внимание и ожидание результата).
Этот путь участвует в обработке зрительной информации на ранних этапах, что приводит к стабилизации зрительной картины (внутренней модели внешнего мира, сохраняющейся благодаря обработке вестибулярной, соматосенсорной и зрительной информации), несмотря на постоянные движения глаз, а также облегчает восприятие зрительной картины и придает различные смыслы в зависимости от модели поведения.
(А) Правое полушарие, вид с медиальной стороны, показан зрительный «Что?» проводящий путь.
Звездочкой отмечена область зрительной идентификации в веретеновидной извилине на нижней поверхности.
Вентральная область 19 увеличена на рис. (Б).
ДМЗ — дополнительная моторная зона.
6. Терминология V1-V5. Специалисты в области исследования зрения используют при обработке корковой зрительной информации следующие обозначения.
V1 соответствует полю 17 по Бродману.
V2 и V3 соответствуют полям Бродмана 18 и 19 соответственно.
V4 включает три группы идентификационных модулей в веретеновидной извилине.
V5 обозначает модули регистрации движений в латеральной затылочной коре (переднелатеральный отдел поля 19 по Бродману) — зрительный путь «Где?».
и) Слуховая кора (поля 41,42 и 22). Первичная слуховая кора расположена в передней поперечной височной извилине Гешля, описанной в отдельной статье на сайте. Извилина Гешля соответствует полям 41 и 42 на верхней поверхности верхней височной извилины; большая часть информации от медиального коленчатого тела направляется к полю 41. Колончатая организация коры предположительно представлена в виде одночастотных полос, т.е. каждая из полос соответствует определенной частоте тона. Высокие частоты активируют латеральные полосы в извилине Гешля, а низкие частоты — медиальные полосы. В связи с неполным перекрестом центральных слуховых путей в стволе мозга (глава 20) каждое ухо имеет двустороннее представительство в коре. По данным исследований, первичная кора одинаково отвечает на монофоническую стимуляцию каждого слухового пути, однако кора противоположной стороны лучше отвечает на одновременную стимуляцию обоих слуховых путей.
Ассоциативная слуховая кора соответствует полю 22, обрабатывающему речь. Одновременная обработка зрительной и слуховой информации происходит в полимодальной коре, ограниченной верхней височной бороздой (соединение полей 21 и 22).
Удаление слуховой коры с одной стороны (например, при опухолевом процессе) не приводит к заметному ухудшению звуковосприятия. Единственный значительный дефект — потеря стереоакузии: при исследовании у пациента возникают трудности с определением локализации и расстояния от источника звука.