После проникновения вириона тогавируса в клетку синтезируется вирусная полимераза (РНК-зависимая РНК-полимераза), катализирующая образование -РНК на матрице +РНК. Образовавшеся цепи -РНК служат матрицей для синтеза двух типов +РНК (полная и короткая нити). Каждая +РНК транслируется в большой полипептид, подвергающийся последовательному расщепленния и процессингу. Полная нить (42 S) служит шаблоном для синтеза вирусных полипетидов или идёт на построение геномов дочерних популяций вируса; короткая нить (26 S) кодирует белки капсида и два оболочечных белка.
Последние гликолизируются, расщепляются и встраиваются в клеточную мембрану, модифицируя ее. Дочерние вирионы тогавируса почкуются через изменённые участки мембраны с образованием суперкапсида.
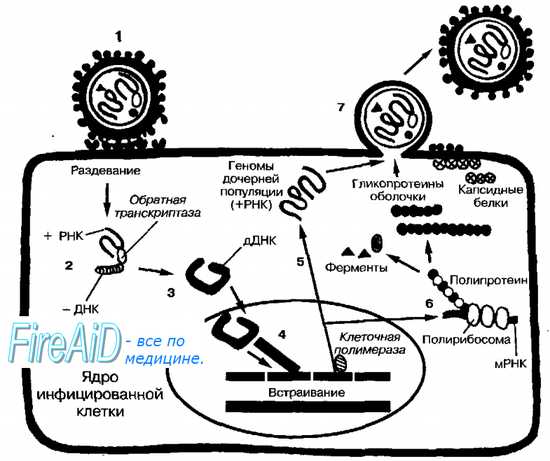
Рис. 5-2. Репродуктивный цикл ретровирусов. Инфекционная вирусная частица (1) проникает в клетку путём слияния с клеточной стенкой после адсорбции на ней. Затем вирус «раздевается», а обратная транскриптаза индуцирует синтез -ДНК, используя в качестве матрицы молекулу +РНК (2) +ДНК копируется с синтезированной молекулы -ДНК, в результате чего образуется двойная цепь ДНК (дДНК) (3). дДНК транспортируется в ядро клетки, где клеточная ДНК подвергается ДНК-сплайсингу (связывание одной молекулы ДНК с другой) с образованием рекомбинантов с вирусной ДНК (4). Интегрированная молекула ДНК транскрибируется клеточной ДНК-зависимой РНК-полимеразой с образованием вирусной +РНК (5). Часть молекул +РНК используется для создания геномов дочерних популяций, другая часть подвергается РНК-сплайсингу {устранение интронов из мРНК-предшественника) с образованием мРНК малого размера для синтеза (через стадию образования полипротеинов структурных белков и ферментов (6). Зрелые дочерние популяции высвобождаются почкованием (7)
Ретровирусы. Репродукция ретровирусов
Репродуктивный цикл +РНК-ретровирусов уникален, так как при его реализации как промежуточный продукт образуются молекулы ДНК (рис. 5-2). Поскольку обратная транскрипция и интеграция вирусного генома предшествуют репликации, то (в противоположность прочих молекулам вирусных +РНК) плюс-молекулы РНК ретровирусов не проявляют инфекционных свойств, а служат матрицей для синтеза молекулы -ДНК РНК-зависимой ДНК-полимеразой (обратной транскриптазой, ревертазой), входящей в состав вирусной частицы. С молекул -ДНК копируются цепи +ДНК. Затем обе цепи соединяются с образованием двойной цепи и транспортируются в ядро клетки. Клеточные эндонуклеазы и лигазы (интегразы) катализируют сплайсинг ДНК клетки с вирусной ДНК. В результате образуются новые, рекомбинантные молекулы ДНК. Вновь образованная ДНК транскрибируется клеточной ДНК-зависимой РНК-полимеразой в +РНК.
Полная копия +РНК способна формировать геномы дочерних популяций, а также транслироваться как мРНК с образованием структурных белков и ферментов, участвующих в сборке вирионов. Эти копии могут быть подвергнуты РНК-сплайсингу — устранению интронов из мРНК-предшественника; в результате образуются малые молекулы мРНК, кодирующие поверхностные, регуляторные и добавочные белки вируса. Самосборка вирусов и созревание вирусных частиц происходит в цитоплазме. Зрелые вирионы выходят из клетки, отделяясь от неё почкованием (высвобождение вируса в окружающую среду может длиться годами).