Катаболизм углеводов у бактерий. Гликолиз. Гликолитический путь окисления. Путь Эмбдена-Мейерхофа-Парнаса. Пентозофосфатный путь окисления. Схема Варбурга-Диккенса-Хореккера-Рэкера.
В качестве источника питания и энергии при выращивании в искусственных условиях бакте рии наиболее часто используют углеводы. Способность утилизировать различные углеводы -важный диагностический признак. Базовый субстрат — глюкоза, которую бактерии разла гают в процессе дыхания либо брожения. Многие реакции катаболизма глюкозы протекают одинаково у аэробных и анаэробных бактерий, в частности три пути превращения гексоз в триозы (гликолиз, пентозофосфатный путь и путь Энтнера-Дудорова).
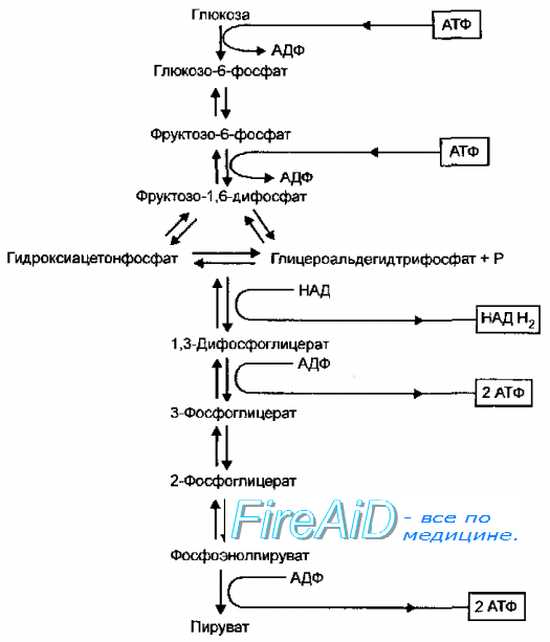
Гликолиз. Гликолитический путь окисления. Путь Эмбдена-Мейерхофа-Парнаса.
Гликолитический путь (гликолиз, путь Эмбдена-Мейерхофа-Парнаса, фруктозо-1,6-дифосфатный путь) доминирует у большинства аэробных и анаэробных микроорганизмов — кишечной палочки, бацилл, пенициллов, дрожжей, стрептомицетов и многих других. Характерная реакция гликолиза — расщепление фруктозо-1,6-дифосфата альдолазой, в результате чего образуется смесь триозофосфатов, состоящая из дигидрооксиацетонфосфата и глицероальдегидтрифосфата, которые затем превращаются в пируват (рис. 4-8). Фермент обратной связи, лимитирующий скорость процесса, — фруктозо-6-фосфат дегидрогеназа (фосфофруктокиназа). В этом процессе образуется 2 моля АТФ и 2 моля восстановленного НАД (НАДН+ на 1 моль глюкозы.
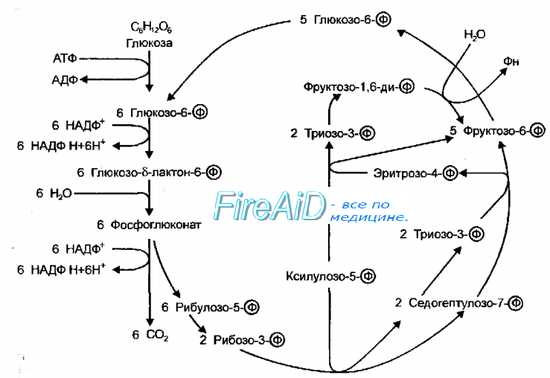
Рис. 4-9. Пентозофосфатный путь окисления. Схема Варбурга-Диккенса-Хореккера-Рэкера.
Пентозофосфатный путь окисления. Схема Варбурга-Диккенса-Хореккера-Рэкера.
Пентозофосфатный путь (схема Варбурга-Диккенса-Хореккера-Рэкера, фосфоглюконатный путь) включает не только образование пентозофосфатов, но и реакции превращения пентозофосфатов в триозо-3-фосфаты (рис. 4-9). В результате изомеризации фруктозо-6-фосфата в глюкозо-6-фосфат и конденсации двух молекул триозо-3-фосфата в гексозофосфат все перечисленные реакции замыкаются в цикл, при одном обороте которого из 3 молекул глюкозо-6-фосфата образуются 2 молекулы фруктозо-6-фосфата, одна молекула триозо-3-фосфата, 3 молекулы С02 и трижды по 2 восстановленных НАДФ (НАДФН+). Последовательность подобных превращений глюкозы у бактерий идентична таковой у высших организмов. Этот путь катаболизма глюкозы следует рассматривать как второстепенный, но имеющий специальное назначение — источник пентоз для синтеза нуклеиновых кислот, а также источник восстановительных эквивалентов НАДФН+.