Fab-фрагменты антитела взаимодействуют с антигенными детерминантами. Аг-связывающий центр комплементарен эпитопу Аг (принцип ключ-замок). Связывание Аг с AT нековалентно и обратимо. Аффинность (сродство) Аг к антитела определяется физико-химическими свойствами взаимодействующих молекул и соотношением концентраций связанных и свободных Аг и антител. На сродство влияют пространственное соответствие взаимодействующих участков молекул, электростатические, гидрофобные взаимодействия и силы Ван дер Ваальса.
Авидность — интегральная характеристика силы связи между Аг и AT, учитывающая взаимодействие всех активных центров с эпитопами Аг.
Валентность антитела — число активных (Аг-связывающих) центров антитела. Молекула полного Ig как минимум двухвалентна. Такие антитела известны как полные антитела; мономеры с меньшей валентностью — неполные антитела.
Полные антитела (в частности, IgM, lgG) вызывают агрегацию Аг, видимую невооружённым глазом (например, РА бактерий).
Неполные антитела содержат один Аг-связывающий центр и, поэтому, одновалентны (например, антитела, вырабатываемые при бруцеллёзе). Второй Аг-связывающий центр у подобных Ig экранирован различными структурами либо обладает низкой авидностью.
Неполные антитела функционально дефектны, так как не способны агрегировать Аг. Неполные AT могут связывать эпитопы Аг, препятствуя контакту с ними полных антител; поэтому их также называют блокирующими антителами.
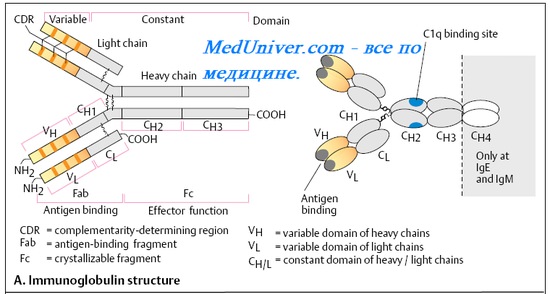
Fc-фрагмент антитела
Константные участки тяжёлых цепей определяют характер взаимодействий антитела с клетками и молекулами иммунной системы, в частности специфичность связывания молекулы Ig с клетками-эффекторами (например, фагоцитами, тучными клетками), несущими на своей поверхности рецепторы к Fc-фрагменту.
Fc-фрагмент определяет также эффекторные функции антитела (например, активацию комплемента). Для реализации этих свойств сразу после связывания Аг Fab-фрагментами происходят конформационные изменения структуры Fc-фрагментов. Пространственно изменённые Fc-фрагменты распознают фагоциты, именно они способствуют фиксации С1а-компонента комплемента и запуску комплементарного каскада по классическому пути. В противном случае ни клетки, ни эффекторные молекулы были бы не в состоянии отличить интактные AT или антитела, связавшие Аг.