Пикорнавирусы. Семейство пикорнавирусов. Виды пикорнавирусов.
Данное семейство включает более 200 вирусов, которые объединены в 6 родов: энтеро-, афто-, кардно-, рино-, гепато- и парэховирусы. Вирусы, вызывающие заболевания человека, входят в 4 рода, а вызывающие заболевания животных - в 6 родов. Важным признаком дифференцирующим родовую принадлежность является стабильность при низком рН. Афтовирусы нестабильны при рН ниже 7,0; риновирусы — при рН ниже 5,0; энтеро-, гепато-, кардио- и парэховирусы стабильны при рН=3,0. 5'-нетранслируемая область генома кардио- и афтовирусов содержит длинный поли (С) участок, отсутствующий у представителей других родов. Афтовирусы уникальны по наличию в геноме трех подобных, но не идентичных участков, кодирующих белок VPg.
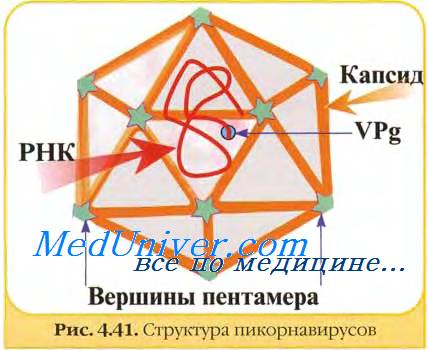
Вирионы представляют собой безоболочечные частицы, округлой формы с гладкой поверхностью диаметром 27 нм.
Геном представлен одной молекулой одноцепочечной (+)РНК размером 7,2—8,4 тн. Геномная РНК полиаденилирована на З'-конце и имеет белок VPg, связанный ковалентно с 5'-концом. Геномная РНК обладает инфекционностью, т.е. функционирует как мРНК, имеет одну открытую рамку считывания и транслируется в полипротеин, который затем расщепляется на 11 индивидуальных белков. Пикорнавирусы содержат 60 копий каждого из 4 капсидных белков: VP1, VP2 и VP3 (м.м. каждого 30000) и VP4 (м.м. 7000-8000) и 1 копию небольшого белка VPg (м.м. варьирует; афтовирусы кодируют 3 варианта VPg). Кроме того, в вирионах многих пикорнавирусов были обнаружены минорные белки, функция которых неизвестна.
Три белка VP1, VP2 и VP3, структурноподобных друг другу, образуют наружную поверхность вириона, а белок VP4 расположен внутри капсида и, вероятно, связан с геномной РНК. Белок VPg участвует в репликации РНК и, вероятно, выполняет сигнальные функции при инкапсидации.
В вирионах полио- и риновирусов, VP1, VP2 и VP3, упакованные вместе, образуют «каньон», обрамляющий 5-гранник. Аминокислоты внутри каньона и особенно в основании, являются вариабельными. Консервативные аминокислоты, расположенные в основании каньона, вероятно, формируют точки прикрепления вирусов к рецепторам на поверхности клетки и, предположительно, защищают их от иммунных механизмов.
В этом отношении риновирусы подобны вирусу ящура, который имеет относительно гладкую поверхность без структуры каньона; места прикрепления к клеточным рецепторам локализованы на верхушке выступов поверхности вириона. Эти участки являются весьма антигенными и определяют серотиповую специфичность вируса ящура. Установлены общие закономерности в антигенной структуре пикорнавирусов. Инфекция или иммунизация (146 S-частицами, но не 12-14 S-субъединицами) сопровождаются образованием вируснейтрализующих антител (ВНА). Главный иммуногенный сайт локализован в VP1, хотя нейтрализующие эпитопы локализованы в двух других капсидных белках - VP2 и VP3. Обнаружена многокомпонентность эпитопов, учавствующих в нейтрализации вируса. Замена только одной аминокислоты в эпитопе в процессе мутации может привести к изменению антигенной структуры пикорнавирусов. Ренгеноструктурный анализ позволил локализовать нейтрализующие антигенные участки энтеровирусов. У вируса полиомиелита идентифицировано три основных нейтрализующих участка: первый находится в VP1 и включает область аминокислот 89-100; второй - аминокислот 220-222 полипептида VP1 и аминокислот 270 или 164—172 полипептида VP2; третий — области аминокислот 58—60 и 70—74 полипептида VP3 или аминокислот 286—290 полипептида VP1 и 58-59 полипепттида VP3. таким образом, вирус полиомиелита содержит три основных антигенных участка, в которые вовлечены три капсидных белка. Первый нейтрализующий участок - непрерывный (области 89-100 аминокислот VP1), второй и третий — нейтрализующие состоят из двух областей: второй - из участков аминокислот белков VP1, VP2, третий - из участков аминокислот полипептида VP3 или VP1 и VP3.
Антигенные участки, связывающие антитела, не участвуют во взаимодействии полиовируса с клеточными рецепторами. У антигенных вариантов полиовируса устойчивость к нейтрализующим антителам сопровождалась аминокислотными заменами во всех трех наружных капсидных белках. На VP1 полиовируса типа 1 идентифицирован антигенный участок нейтрализации 1В. Он представлен эпитопом, который формируется двумя петлями VP1, а также включает аминокислотные остатки 69—104 и 141 — 152. Моноклональные антитела, реагирующие с VP1 полиовируса типа 1, связывались с областью аминокислот 93—104 указанного полипептида. Антигенные сайты сохраняли иммуногенную активность в инвактивированной вакцине. Протеолитическая активизация VP1 полиовируса повышала его антигенность для мышей. Вирус гепетита А, в отличие от полиовируса, по-видимому, обладает одним доминантным нейтрализующим участком. Хотя у энтеровирусов отсутствует групповой антиген, тем не менее, выявлены перекрестные реакции с антисыворотками на денатурированные антигены.
У энтеровирусов парнокопытных животных, кроме того, обнаружены общие эпитопы в полипептидах VP1. Вирус везикулярной болезни свиней содержит три белка: VP1, VP2 и VP3 с молекулярной массой соответственно - 33, 29 и 32 кД. Белки VP1 и VP2 ответственны за индукцию ВН-антител. Энтеровирусы энцефаломиелита и нефрита птиц антигенно не связаны между собой.
У вируса ящура VP1 является иммунодоминантным полипептидом. Он ответственен за индукцию ВНА, а Т- и В-клеточные эпитопы находятся, вероятно, в целой вирусной частице. Кроме VP1, важные антигенные области находятся в структуре VP2 и VP3. Три капсидных белка, экспонированных на поверхности вируса ящура, формируют четыре антигенных участка. Первый из них представлен аминокислотными остатками 140-160 белка VP1, второй, третий и четвертый соответственно — VP1, VP2 и VP3. На поверхности вируса ящура обнаружено три участка нейтрализации. Один выявлен только на интактных вирионах (140S), второй — на инактивированных вирионах и субединицах (12 S) и третий на 140S, 12S — структурах и полипептиде VP1. Главный антигенный сайт вируса ящура - это участок соответствующий 141 — 160 аминокислотным остаткам белка VP1, образующий петлю G-P, выступающую на поверхности вириона. За антигенную специфичность вируса ответственны два вариабельных участка, включающие аминокислотные остатки 42—60 и 134—158. Основным результатом «иммунологических» мутаций вируса ящура во время эпизоотических вспышек (антигенный дрейф) является изменение аминокислотной последовательности в VP1.
Эпитопы, расположенные в VP1, в отличие от эпитопов, расположенных в VP2, чувствительны к трипсину.
Субвирусные частицы 12S вируса ящура содержат высококонсервативный белок, который выявляется моноклональными антителами одной специфичности у шести из семи известных типов вируса. Однако иммунизация ими не сопровождалась образованием ВН-антител. Возможно, в создании специфической защиты существенную роль играют Т-хелперы и антителозависимые факторы иммунитета. Введение инактивированной противоящурной вакцины крупному рогатому скоту сопровождается образованием антител к капсидным белкам и полимеразе ЗД, тогда как при репликации инфекционного вируса дополнительно образуются антитела к неструктурным вирусным белкам (2В, 2С, ЗАВ 1 и/или ЗС). Репликация инфекционного вируса вызывает синтез последних независимо от предварительной вакцинации или клинического проявления заболевания. Данное явление дает возможность выявлять репликацию вируса ящура и различать иммунных и инфицированных животных.
Нейтрализующий эпитоп риновируса человека типа 2 включает аминокислотные остатки 153-164 2Р2. Зрелые вирионы пикорнавирусов по антигенности значительно превосходят собственные субвирусные частицы, образуемые как в процессе синтеза и морфогенеза вирионов, так и при их дезинтеграции. Это, по-видимому, в основном связано с конформационными изменениями структурных полипептидов. При денатурации различными факторами полиовирус, например, может потерять сердцевину. Такая ДС-антигенная конверсия сопровождается потерей способности индуцировать образование ВН-антител. У вируса ящура образование ВН-антител практически вызывали только полные вирионные (140S частицы), а не их компоненты. На этом основании об иммуногенности инактивированных вакцин против данных заболеваний можно судить по концентрации соответственно D- или 140S антигена.