Размножение бактериофагов. Адсорбция бактериофага. Инъекция фага. Репродукция бактериофага. Выход дочерних популяций бактериофага. Негативные колонии бактериофага.
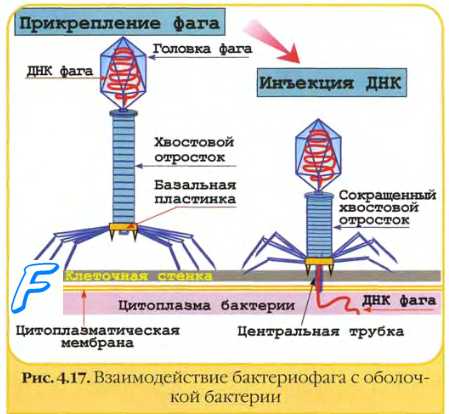
Взаимодействие бактериофагов с клеткой специфично, так как они, как правило, инфицируют бактерии только определённого вида (рис. 5-11). Подобно вирусам животных, репродуктивный цикл литических бактериофагов включает адсорбцию свободного фага на клетке, инъекцию ДНК, репродукцию фага, выход дочерних популяций.
Адсорбция бактериофага
Прикрепление фага к бактерии происходит при помощи поверхностных структур бактериальной стенки, служащих рецепторами для вирусов. Например, рецепторы для фагов ТЗ, Т4 и расположены в липополисахаридном слое, для Т2 и Т6 — в наружной мембране. На бактериях клеточной оболочки (протопласты, L-формы) бактериофаги не адсорбируются. Некоторые фаги в качестве рецепторов используют F-пили. Помимо рецепторов, адсорбция фага зависит от рН среды, температуры, наличия катионов и некоторых соединений (например, триптофана для Т2-фага). При избытке фага на одной клетке может адсорбироваться до 200-300 вирусных частиц.
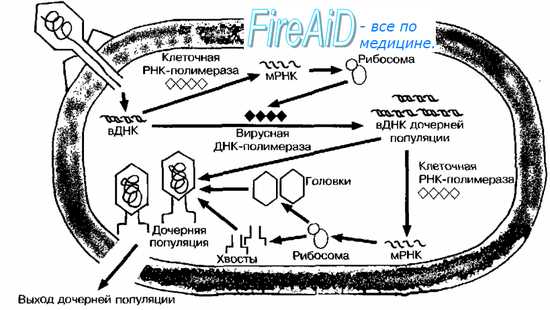
Рис. 5-11. Литическое взаимодействие фага с бактериальной клеткой. Бактериофаг вводит в вирусную ДНК (вДНК) в цитоплазму бактериальной клетки. Клеточные РНК-полимеразы транскрибируют ДНК в мРНК, транслирующуюся на рибосомах. В результате осуществляется синтез вирусной полимеразы и других ранних вирусных белков. Вирусная полимераза участвует в образовании вДНК дочерних популяций. Часть образовавшейся вДНК используется как матрица для синтеза белков головок и хвостов. После присоединения вДНК последние образуют дочернюю популяцию фагов.
Инъекция бактериофага
После адсорбции происходит ферментативное расщепление клеточной стенки лизоцимом, находящимся в дистальной части отростка. Базальная пластина хвоста лизирует прилегающий фрагмент клеточной стенки, выделяя присутствующий в отростке лизоцим. Одновременно в чехле высвобождаются ионы Са2+, активизирующие АТФазу, что вызывает сокращение чехла и вталкивание стержня хвоста через ЦПМ в клетку. Затем вирусная ДНК впрыскивается в цитоплазму (внедрение вирусной ДНК). Поскольку диаметр канала лишь немного превышает диаметр молекулы ДНК (около 20 нм), то ДНК способна попадать в цитоплазму только в форме нити.
Репродукция бактериофага
Проникнув в клетку, ДНК фага «исчезает»; уже через несколько минут обнаружить вирус не удаётся. В этот, так называемый скрытый период (эклипс) вирус берёт на себя генетическое управление клеткой, осуществляя полный цикл репродукции фага. К его окончанию составляющие фага соединяются в зрелый вирион.
Синтез фаговых белков. В первую очередь синтезируются ферменты, необходимые для образования копий фаговой ДНК. К ним относятся ДНК-полимераза, киназы (для образования нуклеозидтрифосфатов) и тимидилат синтетаза. Они появляются в клетке через 5-7 мин после её заражения. Клеточная РНК-полимераза транскрибирует вирусную ДНК в мРНК, которая транслируется бактериальными рибосомами в «ранние» белки фага, включая вирусную РНК-полимеразу и белки, способные посредством различных механизмов ограничивать экспрессию бактериальных генов. Вирусная РНК-полимераза осуществляет транскрипцию «поздних» белков (например, белков оболочки и эндолизина), необходимых для сборки фаговых частиц дочернего поколения. Некоторые вирусы расщепляют ДНК клетки-хозяина до нуклеотидов, чтобы использовать их для синтеза собственных нуклеиновых кислот.
Репликация нуклеиновых кислот реализуется за счёт активности вновь синтезированных вирусных ДНК-полимераз, производящих множественные копии вирусных нуклеиновых кислот.
Выход дочерних популяций бактериофага
Вновь синтезированные белки формируют в цитоплазме пул предшественников, входящих в состав головок и хвостов дочерних вирусных частиц. Другой пул содержит ДНК потомства. Специальные аффинные области в вирусной ДНК индуцируют объединение предшественников головок вокруг агрегатов нуклеиновой кислоты и образование ДНК-содержащих головок. Заполненная головка затем взаимодействует с хвостовой частью, образуя функциональный фаг. Весь процесс (от адсорбции до появления вновь синтезированных вирусов) занимает около 40 мин. После образования потомства («урожай», или выход фага, составляет 10-200 из одной инфицирующей частицы) клетка хозяина лизируется, высвобождая дочернюю популяцию. В разрушении клеточной стенки участвуют различные факторы: фаговый лизоцим, увеличенное внутриклеточное давление. Вирус, по-видимому, также стимулирует образование аутолизинов либо блокирует механизмы, регулирующие их синтез (подобные литические факторы выявлены в фаголизатах многих бактерий).
Негативные колонии бактериофага
Размножение фагов в бактериальных культурах, засеянных сплошным «газоном» на твёрдых средах, сопровождается лизисом бактерий и образованием зон просветления — «стерильных пятен». Для их обозначения предложен термин «негативные колонии бактериофага». У разных фагов они имеют строго определённые размеры и форму (например, звёздчатую у дизентерийных фагов). При заражении бульонных культур литическим фагом наблюдают просветление среды. Способность образовывать негативные колонии в культурах чувствительных бактерий в микробиологической диагностике возбудителей инфекционных болезней известна как фагодиагностика