Механизм работы молекулярных моторов (моторных белков)
• Для осуществления движения моторные белки используют энергию гидролиза АТФ
• Прочность связывания домена головки мотора с микротрубочкой определяется нуклеотидом, ассоциированным с головкой. Это может быть АТФ или АДФ. Связывание может происходит и без участия нуклеотида
• При гидролизе АТФ меняется также форма головки. Это изменение формы амплифицируется, что обеспечивает более выраженное движение мотора
• Циклы гидролиза АТФ и высвобождения нуклеотида сопряжены с присоединением к микротрубочке и с изменениями формы домена головки мотора. Используя этот механизм, мотор движется по микротрубочке, делая один шаг на каждый цикл гидролиза АТФ
В качестве топлива молекулярные моторы используют АТФ, но каким образом моторные белки превращают химическую энергию АТФ в механическую работу? В данном разделе мы рассмотрим, как происходит движение моторных белков по микротрубочкам. Мы бы очень мало знали об этом, если бы не имели возможности наблюдать за движением моторов. Здесь мы не будем подробно рассматривать, каким образом исследуют эти процессы, однако интересующиеся читатели могут найти всю информацию в отдельных статьях на сайте (рекомендуем пользоваться формой поиска на главной странице сайта).
Основное требование, предъявляемое к мотору, заключается в том, чтобы за время между присоединением АТФ и образованием АДФ могли пройти очень существенные конформационные перестройки. Это достигается за счет изменений моторного домена и примыкающих к нему участков молекулы и напоминает процесс управления движением наших конечностей. В обоих случаях небольшие локальные изменения формы или положения усиливаются и приводят к гораздо более выраженным изменениям в другом месте. Например, относительно небольшое сокращение мышцы бедра при ходьбе вызывает перемещение голени вверх и вперед, и, таким образом, незначительные изменения длины мышцы усиливаются, вызывая гораздо более выраженные изменения в положении ноги. В случае моторных белков, в результате гидролиза АТФ до АДФ в области связывания нуклеотида (нук-леотид-связывающий карман) происходят небольшие изменения формы головки моторного домена. Эти изменения усиливаются, проявляясь в другой части молекулы в перемещении вперед одной из головок
Так же как и пешеходы, молекулярные моторы должны быть способны открепляться от поверхности, на которой они находятся; в противном случае ни те, ни другие не смогут продвигаться дальше. Так же как и пешеход, для того, чтобы шагнуть вперед должен поднять ногу, белковый мотор должен открепиться от микротрубочки для дальнейшего продвижения. Насколько прочно мотор связывается с микротрубочкой, зависит от содержимого нуклеотид-связывающего кармана. Он может не содержать нуклеотидов, содержать АТФ или АДФ. Для кинезинов связывание с микротрубочками оказывается наиболее прочным, когда в области кармана находится связанный АТФ.
Связь мотора с микротрубочкой регулируется за счет гидролиза АТФ и при высвобождении нуклеотида. Это происходит за счет изменения прочности связи кинезина с микротрубочкой. Поскольку при гидролизе АТФ также изменяется форма головки моторного домена, процессы связывания нуклеотида, его гидролиза и высвобождения обеспечивают координацию между формой мотора и его связыванием с микротрубочкой. Это позволяет мотору сделать один «шаг» на каждую молекулу гидролизованного АТФ, т. е. осуществить один цикл: связывание с микротрубочкой, изменение конформации и высвобождение нуклеотида.
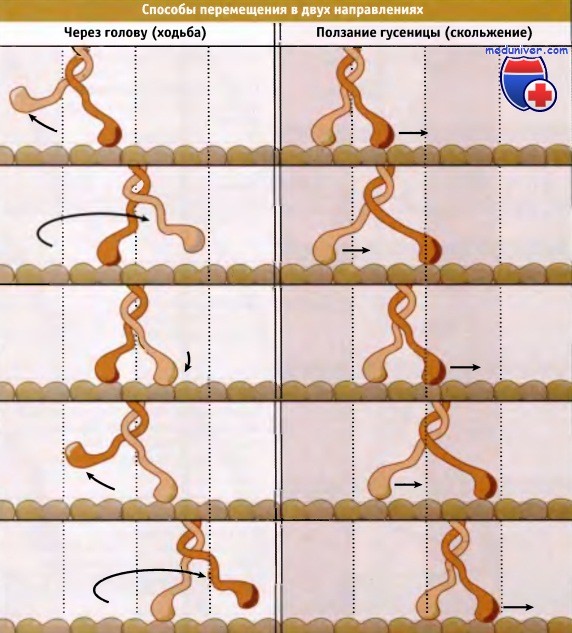
Для мотора с двумя головками можно представить два различных способа перемещения вдоль микротрубочки. Мотор может шагать «через голову», как это представлено на рисунке ниже. При этом, при каждом шаге вперед, головка, расположенная сзади, проходит мимо передней головки. Такой способ напоминает движения, которые мы совершаем при ходьбе, с каждым шагом перенося вперед ногу. Другой способ предполагает, что движение доменов мотора напоминает ползущую гусеницу. Головка, расположенная сзади, подтягивается по направлению к передней, затем передняя начинает двигаться вперед и цикл повторяется. Все известные моторы с двумя головками перемещаются по микротрубочкам, шагая «через голову». Иными словами, можно представить, что кинезин и другие моторы «прогуливаются» по микротрубочке.
Два способа перемещения мотора с двумя головками по микротрубочке.
При согласованных движениях двух головок кинезин перемещается как ползущая гусеница (справа).
При этом головка, обозначенная красным цветом, продвигается вперед, а помеченная оранжевым к ней подтягивается, и весь цикл повторяется.
Оранжевая головка никогда не занимает место впереди красной. Хотя существование этого способа вполне вероятно, такой мотор пока не найден.
Все известные моторы с двумя головками перемещаются по шагающему механизму: две головки перемещаются одна за другой, попеременно становясь ведущими (слева).
Важную роль в перемещении кинезина играет малый домен. Он называется шейным линкером и представляет собой последовательность, состоящую из 15 аминокислот, которая находится между головкой и спирализованной ножкой. Шейный линкер является частью молекулы кинезина, которая усиливает небольшие изменения структуры нуклеотид-связывающего кармана для того, чтобы обеспечить кинезину возможность делать длинные шаги. Усиление достигается за счет качательных движений шейных линкеров, которые можно рассматривать как «ноги» молекулы кинезина. Как мы вскоре увидим, перемещение кинезина обеспечивается АТФазным циклом обеих головок, контролирующих движение шейных линкеров.
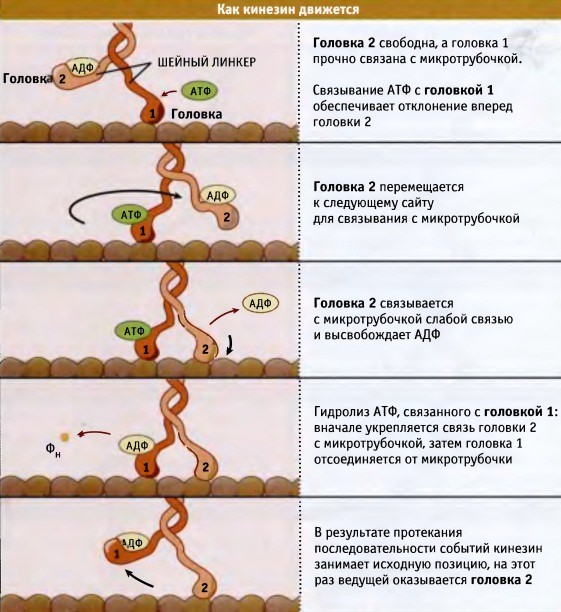
При продвижении кинезина по микротрубочке обе головки работают в тандеме. Изменения, происходящие в одной из головок, часто являются результатом изменений в другой. Чтобы понять, каким образом цикл гидролиза АТФ обеспечивает движение мотора по микротрубочке, начнем с момента прикрепления кинезина. Одна из головок прочно зафиксирована на микротрубочке, и ее связывающий карман не содержит нуклеотидов. За головкой расположен шейный линкер. Связывающий карман второй головки содержит АДФ, и сама головка свободно расположена вдоль микротрубочки. Кинезин готов сделать первый шаг, и между обеими головками начинают разворачиваться события, координирующие их дальнейшие движения. При связывании АТФ с передней головкой (головка 1) ее шейный линкер отклоняется вперед по направлению к плюс-концу микротрубочки.
Это движение приводит к тому, что головка 1 перемещает головку 2 в лидирующее положение, и она располагается над следующим сайтом связывания на микротрубочке. Затем головка связывается с сайтом и отщепляет АДФ. Гидролиз АТФ, связанного с головкой 1, приводит к упрочению взаимодействия между головкой 2 и микротрубочкой. Кинезин переходит в промежуточное состояние, при котором обе головки оказываются прочно связанными с микротрубочкой. Как только головка 2, которая теперь занимает лидирующее положение, прочно связалась с микротрубочкой, головка 1 высвобождает фосфатную группу, образующуюся при гидролизе АТФ. Этот процесс приводит к отщеплению головки 1 от микротрубочки и вызывает конформационные изменения в головке 2, в результате которых повторно открывается активный сайт. Вся цепь описанных событий возвращает кинезин в исходное состояние лишь с той разницей, что впереди теперь находится головка 2, и молекула кинезина расположена на 8 нм ближе к плюс-концу микротрубочки.
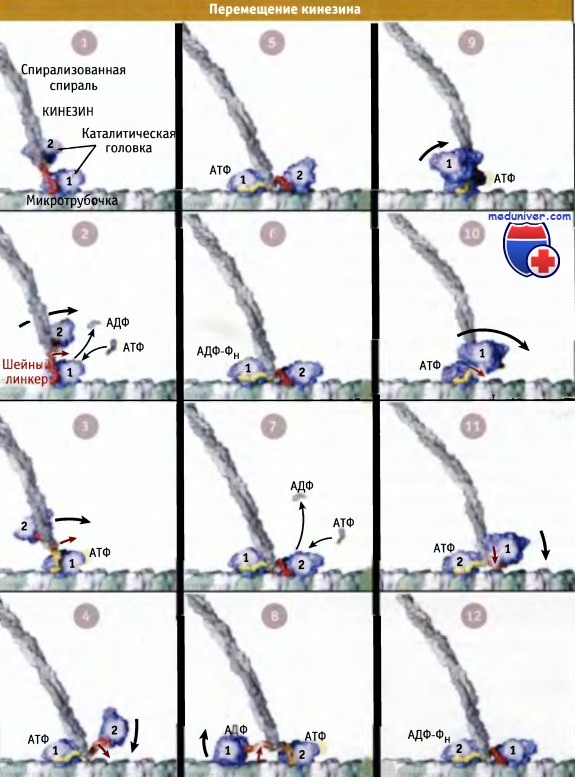
Когда головка 2 связывает АТФ, наступает второй цикл, и затем две головки будут меняться ролями в течение сотен или тысяч циклов, делая каждый раз по шагу и продвигая мотор вдоль микротрубочки по направлению к плюс-концу. На рисунке ниже представлена серия анимированных изображений молекулы кинезина, которая делает несколько последовательных шагов. Схема иллюстрирует, каким образом координация процессов с участием обеих головок и обмен ролями между ними после каждого шага позволяет мотору двигаться по микротрубочке.
Последовательность процессов, которые происходят при выполнении кинезином каждого шага, напоминают действия канатоходца, шагающего по канату, закрепленному высоко над землей. Вначале кинезин, стоя на одной ноге, сохраняет равновесие и находится в устойчивом положении. Затем он переносит другую ногу вперед (изменение положения шейного линкера) и осторожно нащупывает ею канат (предварительное связывание передней головки). Только когда кинезин убедится в том, что передняя нога надежно ориентирована на канате, он переносит на нее свой вес (укрепление связывания передней головки). Затем кинезин высвобождает вторую ногу и отрывает ее от каната, принимая положение для следующего шага. Если кинезин, так же как и канатоходец, переставит вторую ногу, прочно не укрепив первую, прогулка закончена. Канатоходец оказывается на земле, а белок далеко от микротрубочки.
Если бы кинезин часто освобождал вторую ногу, не дожидаясь, пока передняя нога прочно укрепилась, то постоянное движение на большие расстояния стало бы невозможным.
Механизм перемещения кинезина предполагает существование строгой скоординированности в работе обеих головок Каким образом это достигается? Как одна головка «узнает», что делает другая? По-видимому, сообщение между головками осуществляется при помощи шейных линкеров. Когда обе головки кинезина прочно связаны с микротрубочкой, шейные линкеры выпрямлены и находятся под механическим напряжением. Очевидно, это позволяет головкам сообщаться друг с другом и координировать свою деятельность (например, АТФазный цикл). Наличие механического напряжения указывает одной головке, что другая прочно связана с микротрубочкой. Поэтому одна головка может безопасно отсоединиться от микротрубочки. За счет механического напряжения, вероятно, осуществляется координация деятельности головок, в результате чего устанавливается, насколько быстро проходят различные этапы АТФазного цикла.
Например, если бы механическое напряжение вызывало значительное ускорение высвобождения фосфата, то ведомая головка не откреплялась бы от микротрубочки до момента прочного связывания ведущей головки.
Подвижность динеинов также связана с усилением изменений конформации, однако структурные изменения происходят на еще большем расстоянии. Кинезин и динеин продвигаются по микротрубочке 8-нм шагами, что эквивалентно длине одного тубулинового гетеродимера. Кинезин «осторожно» продвигается по протофиламенту, ступая с одного гетеродимера на следующий. Для сравнения, динеин, при перемещении к минус-концу микротрубочки, «бредет» как пешеход, случайно наступая между протофиламентами.
Механизм движения кинезинов дает им возможность постоянно продвигаться по микротрубочке (т. е. их движение является высоко «процессивным»). Например, в экспериментах in vitro один кинезиновый мотор с присоединенными к нему стеклянными шариками (выполняющими роль удобного карго) и содержащий две головки способен сделать сотни и тысячи шагов вдоль микротрубочки и, не отсоединяясь от нее, переместить шарики на значительное расстояние. Способность одного кинезинового мотора перемещать по микротрубочке карго на большие расстояния связана с тем, что каждая головка мотора примерно половину времени расходует на связывание с микротрубочкой, и активность двух головок скоординирована таким образом, что, по крайней мере, одна из них всегда находится в связанном состоянии. Существование моторов, у которых одна из двух головок всегда связана с микротрубочкой, характерно для индивидуально функционирующих или небольшой группы моторов, например, транспортирующих везикулы. Работая по такому механизму, они способны транспортировать карго на большие расстояния.
Существуют и такие моторы, у которых одна головка не всегда связана с микротрубочкой, что позволяет им вместе с карго быстро терять контакт с ее поверхностью. Моторы, которые функционируют в больших группах, например присутствующие в жгутиках, гораздо меньше времени находятся в связанном с микротрубочкой состоянии, чем моторы, переносящие везикулы. Некоторые головки могут находиться в связанном состоянии и генерировать силу в структуре, содержащей динеины (в жгутиках). Моторы, завершившие свою функцию, быстро покидают микротрубочку, чтобы не мешать работать на ней другим активным моторам.
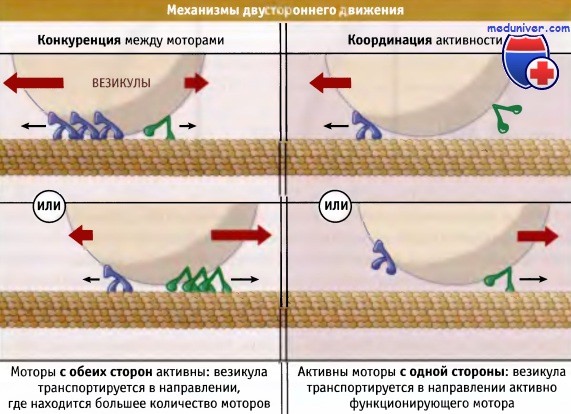
В клетке многие органеллы движутся в двух направлениях: вначале органелла движется в одном направлении вдоль микротрубочки, затем она поворачивается и движется в противоположном направлении. Эти органеллы связываются с динеинами и с представителями группы кинезинов. При этом возникает вопрос, что обеспечивает их продолжительное движение в одном направлении? Для объяснения двустороннего движения органелл предложены две модели, представленные на рисунке ниже. Моторы с противоположно направленной полярностью могут конкурировать между собой. Поскольку всегда активны оба типа моторов, то выигрывает тот, который способен генерировать более сильное усилие (поскольку присутствует в большем количестве). Наоборот, активность моторов может быть скоординирована таким образом, что один их набор выключен, а другой функционирует. Вероятно, в клетке работает второй механизм, однако неясно, каким образом активность моторов скоординирована на поверхности везикулы.
Последовательность процессов, приводящих к перемещению кинезина по микротрубочке.
Несколько раз в ответ на изменения в одной головке, в другой также происходят изменения.
Заметьте, что по меньшей мере одна из головок кинезина всегда прочно связана с микротрубочкой.
Кадры видеоанимации, показывающие скоординированное движение двух головок кинезина по микротрубочке.
Две головки (обозначены 1 и 2) выделены синим цветом, суперспирализованная область,
серым, и область шейного линкера, желтым (при направлении вперед) и красным (при направлении назад).
Для простоты показан только один протофиламент микротрубочки.
а-, b-субъединицы тубулина выделены белым и зеленым цветами соответственно, и плюс-конец расположен справа.
Возможные механизмы двустороннего движения карго по микротрубочке.
В каждом случае оба мотора, транспортирующие карго в плюс- и минус-направлении, связаны с поверхностью везикулы.
На левом рисунке активны оба типа моторов, и мотор, способный развивать большее усилие
(вероятно из-за того, что присутствует в большем количестве копий), определяет направление движения везикулы.
На правом рисунке деятельность моторов скоординирована таким образом, что в каждый момент времени активен мотор, перемещающий карго только в одном направлении.
Имеющиеся данные позволяют считать, что клетки используют механизм, представленный на правом рисунке, однако неясно, каким образом скоординирована деятельность моторов.