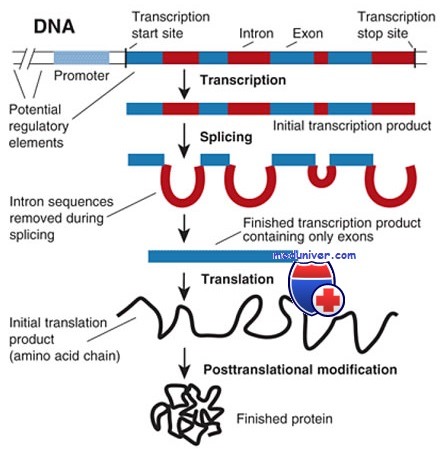
Для генов, кодирующих белки, движение информации от гена до полипептида включает несколько шагов. Инициация транскрипции гена происходит под влиянием промоторов и других управляющих элементов, а также специфических белков, известных как факторы транскрипции, взаимодействующих с определенными последовательностями в пределах управляющих областей гена и определяющих пространственную и временную схему экспрессии гена. Транскрипция гена начинается со «стартовой» точки в хромосомной ДНК в начале 5'-транскрибируемой, но не транслируемой области.
Процесс транскрипции идет непрерывно по ходу кодирующей последовательности вдоль хромосомы, проходя от нескольких сотен пар оснований до более миллиона пар, захватывая как интроны, так и экзоны, и завершаясь на конце кодирующей последовательности. После модификации обоих 5' и З'-концов первичной копии РНК части, соответствующие нитронам, удаляются, а сегменты, соответствующие экзонам, сращиваются вместе.
После сплайсинга (сращивания) РНК результирующая мРНК (содержащая центральный сегмент, соответствующий кодирующей части гена), перемещается из ядра в цитоплазму клетки, где мРНК транслируется в аминокислотную последовательность закодированного полипептида. Каждая составляющая этого сложного пути подвержена ошибкам и мутациям, которые создают помехи и вызывают множество наследственных заболеваний.
Транскрипция
Транскрипция белок-кодирующего гена РНК-полимеразой-II (одна из нескольких классов РНК-полимераз) начинается в стартовом сайте транскрипции, в точке 5'-нетранслируемой области, соответствующей 5'-концу конечной РНК. Синтез первичной копии РНК идет по направлению от 5' к З'-концу, поскольку нить считываемого гена, который служит шаблоном для синтеза РНК, действительно считывается в направлении от 3' к 5'-концу в соответствии с направлением фосфатных связей дезоксирибозы.
Поскольку синтезированная РНК соответствует расположению и последовательности нуклеотидов (с заменой урацила на тимин) 5'-3'-нити ДНК, такую нить ДНК часто называют кодирующей или комплементарной ДНК (кДНК). 3'-5'-нить ДНК носит название некодирующей или антисмысловой. Транскрипция осуществляется как для интронных, так и для экзонных частей гена, до позиции в хромосоме, которая записывается на 3'-конец зрелой мРНК. Неизвестно, заканчивается ли транскрипция в определенной точке терминации на 3'-конце.
Затем в области 5'-конца первичной копии РНК происходит кэпирование, а в специфической точке 3'-конца — расщепление. Расщепление заканчивается присоединением к 3'-концевым звеньям множества остатков аденозина — поли-(А), что увеличивает стабильность полученной РНК. Позиция точки полиаденилирования частично определяется последовательностью AAUAAA (или вариантами этой последовательности), обычно обнаруживаемой в 3'-нетранслируемой части копии РНК. Описанные посттрансляционные модификации, как и процесс сплайсинга РНК, происходят в ядре.
Полностью обработанная РНК, теперь называющаяся мРНК, перемещается в цитоплазму, где происходит трансляция.
Трансляция и генетический код
В цитоплазме мРНК транслируется в белок под действием молекул тРНК, специфичной для каждой конкретной аминокислоты. Эти замечательные молекулы, каждая всего от 70 до 100 нуклеотидов длиной, добавляют к растущей полипептидной цепи определенную аминокислоту в соответствии с шаблоном мРНК. Белковый синтез происходит в рибосомах, макромолекулярных комплексах, состоящих из рРНК (кодируемой генами 18S и 28S) и нескольких десятков рибосомальных белков.
Ключ трансляции — код, который связывает специфическую аминокислоту с комбинацией из трех смежных оснований на мРНК. Каждое сочетание трех оснований составляет кодон, специфичный для конкретной аминокислоты. В теории существует почти бесконечное множество вариантов размещения оснований вдоль полинуклеотидной цепи. В каждом положении может быть один из четырех нуклеотидов (А, Т, С или G); таким образом, для трех оснований есть 43 или 64 возможные комбинации триплетов. Эти 64 кодона и составляют генетический код.
Поскольку на 20 аминокислот приходится 64 возможных кодона, некоторые аминокислоты определяются более чем одним кодоном; поэтому генетический код называют вырожденным. Например, основание в третьей позиции триплета часто может быть или пуриновым (А или G), или пиримидиновым (Т или С), а в некоторых случаях любое из четырех оснований не изменяет смысл сообщения. Лейцин и аргинин определяются сразу шестью кодонами. Только метионин и триптофан кодируются единственным, уникальным триплетом. Три кодона называются стоп-кодонами (или нонсенс-кодонами), поскольку они определяют завершение трансляции мРНК.
Трансляция зрелой мРНК всегда начинается с кодона, определяющего метионин. Следовательно, метионин — всегда первая аминокислота каждой полипептидной цепи, хотя обычно он удаляется до завершения синтеза белка. Кодон метионина (или кодон-инициатор, AUG) устанавливает рамку считывания мРНК; каждый последующий кодон считывается поочередно, указывая аминокислотную последовательность белка.
Молекулярные связи между кодонами и аминокислотами обеспечивают специфические молекулы тРНК. Конкретный участок (сайт) на каждой тРНК формирует антикодон из трех оснований, комплементарный (дополнительный) к специфическому кодону на мРНК. Соединение между кодоном и антикодоном приводит соответствующую аминокислоту на следующую позицию в рибосоме для присоединения с образованием пептидной связи к карбоксильному концу растущей полипептидной цепи. Рибосома затем скользит вдоль мРНК точно на три основания, захватывая следующий кодон для опознавания другой тРНК со следующей аминокислотой.
Таким образом, белки синтезируются, начиная от аминогруппы к карбоксильной группе, что соответствует трансляции мРНК в направлении от 5'-конца к 3'-концу.
Как упоминалось ранее, трансляция заканчивается, когда в той же рамке считывания, что и кодон-инициатор, встречается стоп-кодон (UGA, UAA или UAG). Стоп-кодоны в любой из других неиспользованных рамок считывания не читаются и, следовательно, не оказывают эффекта на трансляцию. Завершенный полипептид отделяется от рибосомы, и она становится доступной для начала синтеза другого белка.
Посттрансляционные события - процессинг
Множество белков проходят существенную посттрансляционную модификацию. Полипептидная цепь, первичный продукт трансляции, скручивается и складывается в специфическую трехмерную структуру, определяемую аминокислотной последовательностью цепи.
Две и более полипептидные цепи, продукты одного или различных генов, могут объединяться, формируя один готовый белковый комплекс. Например, две цепи b-глобина и две цепи а-глобина нековалентно объединяются, формируя тетрамер молекулы гемоглобина. Белковые продукты также могут быть модифицированы химически, например добавлением в специфических местах метильных или фосфатных групп или углеводов.
Такие модификации могут иметь значимое влияние на функцию или количество модифицированного белка. Другие модификации могут включать расщепление белка с потерей специфических аминокислотных последовательностей после того, как они выполнили свою функцию, направив белок в правильную позицию в пределах клетки (например, белки, которые функционируют в пределах ядра или митохондрий) или разделение белковых молекул на меньшие полипептидные цепи.
Например, две цепи, формирующие готовый инсулин, содержащие одна 21, а вторая 30 аминокислот, первоначально представляют собой части проинсулина — первичного продукта трансляции из 82 аминокислот.
Транскрипция митохондриального генома
В предшествующих разделах описаны основы экспрессии генов, содержащихся в ядерном геноме. Митохондриальный геном имеет отличающуюся систему транскрипции и белкового синтеза. Для транскрипции митохондриального генома используется специализированная РНК-полимераза, закодированная в ядерном геноме, содержащая две взаимосвязанные последовательности промотора, для каждой нити кольцевой митохондриальной хромосомы. Каждая нить транскрибируется полностью, а полученные копии затем обрабатываются, порождая различные митохондриальные мРНК, тРНК и рРНК.