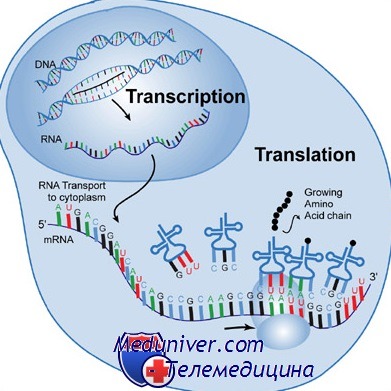
Большинство процессов метаболизма в организме катализируются белковыми ферментами. Кроме того, белки — основные структурные компоненты тела человека. Аминокислотные последовательности всех белков зашифрованы в ДНК, а процесс превращения закодированной информации в сам белок включает её транскрипцию на гяРНК, процессинг на иРНК, трансляцию на полипептид и окончательную сборку белка.
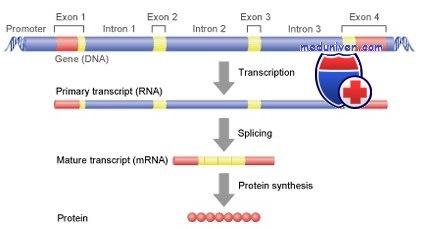
Строение гена
В отличие от прокариот у эукариот большинство генов имеют участок ДНК, который прерывает кодирующую последовательность. Данные некодирующие фрагменты называют нитронами, в то время как другие, кодирующие участки— экзонами. У обеих групп после кодирующего участка присутствуют лидерная и трейлерная последовательности, а также ряд последовательностей, контролирующих процесс транскрипции.
Гены, которые кодируют белок, называют «структурными генами», их транскрипция происходит при участии РНК-полимеразы II. Участок, расположенный непосредственно перед кодирующей последовательностью (в направлении от 3'- к 5'-концу), называют промотором. Он служит для связывания с факторами транскрипции, которые указывают, где РНКаза II должна начать своё действие.
Среди белков различают белки «домашнего хозяйства», которые присутствуют во всех клетках организма, а также белки «роскоши», осуществляющие специальные функции. В состав промоторов генов, кодирующих белки «роскоши», входит «бокс TATA», имеющий последовательность наподобие 5'-ТАТААА-3', повторяющуюся на протяжении примерно 25 пар нуклеотидов и предшествующую участку начала транскрипции.
Гены, кодирующие белки «домашнего хозяйства», имеют на подобных участках один или несколько «боксов ГЦ», составляющих последовательность наподобие 5'-ГГГГДТГ-3. Другой распространённый промоторный участок— «бокс ЦААТ» (например, 5'-ЦЦААТ-3') длиной до 80 пар нуклеотидов, имеющий энхансерные и сайленсерные последовательности на некотором расстоянии от него, которые связывают факторы контроля, взаимодействующие с промотором, образуя петлю ДНК.
Некоторые гены «роскоши» также имеют дополнительные специфические факторы контроля.
Фрагмент, расположенный ниже участка начала транскрипции (5'—3'), называют лидерной последовательностью, он не транслируется. Затем следует кодирующий участок, обычно прерываемый одним или несколькими интронами, а после — некодирующий трейлерный (концевой) участок, на конце которого участок полиаденилирования (поли-А-сайт), имеющий вариабельную последовательность наподобие 5'-ААТАА-3' (5'-ААУАА-3' на РНК-транскрипте) длиной 10-30 пар нуклеотидов в направлении 3'—5'.
Интроны начинаются последовательностью ГТА(/Г)ГАГТ и заканчиваются серией из Ц- или Т-оснований, предшествующих АГ. Для удаления интрона значение имеют первые основания Г и Т (Г и У в гяРНК) и последние АГ, а также остаток аденина в составе последовательности ближе к 5'-концу. Участок, находящийся ближе к 5'-концу, известен под названием донор, ближе к З'-концу — акцептор, а остаток аденина называют участком ветви.
У прокариот транскрипция останавливается на особом участке, состоящем из инвертированного повтора трейлера и ряда Т-остатков. К прекращению транскрипции приводит появление петли-«шпильки», образованной путём спаривания оснований в копии иРНК. Подобная структура существует и в трейлерах гистонных генов. У эукариот не обнаружено общего сигнального участка терминации транскрипции.
Транскрипция при синтезе иРНК
Сигналом к началу транскрипции служит комплекс белков-факторов транскрипции, находящийся в промоторе. Молекула РНКазы II связывается с данным транскрипционным комплексом и разрывает двойную спираль. После этого комплекс, уже имеющий в своём составе фермент, движется подобно «застёжке на молнии» в направлении 5'—3', вызывая разматывание и разделение цепи в месте, где он проходит, а затем восстанавливая структуру двойной спирали сразу после прохождения участка.
Таким образом формируется транскрипционное вздутие. Как только он достигает участка начала транскрипции, происходит отщепление одного из факторов транскрипции и присоединение другого, после чего начинается процесс синтеза РНК.
Используя в качестве матрицы цепь в направлении 3'—5' (слева направо), РНКаза II поочерёдно захватывает рибонуклеотиды и соединяет их друг с другом, образуя комплементарную последовательность РНК, ориентированную в обратном направлении (то есть от 5' к 3').
Другими словами, используя правила комплементарного спаривания оснований при взаимодействии с матричной цепью, РНКаза создаёт точную РНК-копию кодирующей цепи. Фермент транскрибирует лидерный и трейлерный участки, экзоны, интроны и (по всей видимости, напрасно) продвигается дальше в направлении 5'—3'.
Факторы транскрипции при синтезе иРНК
Факторы транскрипции — белки, прикрепляющиеся к промоторной последовательности и запускающие процесс транскрипции. В их состав обычно входят активационный домен и ДНК-связывающий домен. Активационные домены богаты глутаматом, а также аспартатом или пролином, которые облегчают формирование транскрипционного комплекса. Кроме того, различают четыре типа ДНК-связывающих доменов.
• Лейциновая «молния» представляет собой а-спираль, состоящую из аминокислотных остатков, каждый седьмой из которых представлен лейцином, что в свою очередь соответствует каждому второму повороту двойной цепи ДНК. Это позволяет парам оснований сцепляться и образовывать расходящийся участок на конце, который предположительно сжимает нить ДНК наподобие прищепки.
• Спираль-петля-спираль состоит из двух белковых а-спиралей, которые соединены длинной, гибкой петлёй, позволяющей параллельно упаковывать их близко друг к другу. Считают, что данная структура осуществляет контроль процесса транскрипции путём блокирования других регуляторных белков гена.
• Спираль-поворот-спираль состоит из двух коротких а-спиралей, разделённых аминокислотной последовательностью, слишком короткой, чтобы позволить им лежать в одной плоскости. Этот фрагмент — характерный признак гомеобокса (см. главу 12).
• Цинковый палец — структура, напоминающая по строению палец, включающая около 23 аминокислот, удерживаемых четырёхвалентным ионом цинка, который находится в основании «пальца» и обычно связан с четырьмя основаниями цистеина либо двумя — цистеина, двумя — гистидина.
Процессинг РНК
Для того чтобы только что синтезированные гяРНК стали кодирующими матрицами для последующей трансляции и образования полипептидов, они претерпевают ковалентное видоизменение. При этом вначале к 5'-концу в обратном направлении прикрепляется 7-метил-ГТФ (кэп). Как только на цепи гяРНК возникает участок полиаденилирования, она в этом месте расщепляется, а затем при помощи полиА-полимеразы происходит присоединение 100—200 остатков адениловой кислоты и таким образом формируется поли-А-хвост (полнаденильный хвост).
Наряду с кэпом поли-А-хвост предположительно защищает молекулу от разрушения экзонуклеазами, служит так называемым паспортом, необходимым для её попадания в цитоплазму, а позже становится сигнальным участком для рибосомы, указывающим на возможность начала трансляции.
Молекула гяРНК в среднем содержит около 7000 нуклеотидов, количество которых в иРНК сокращается до 1200 путём удаления примерно 50 интронов. Характерная особенность гистонных генов — отсутствие интронов.
Рибонуклеиновые комплексы, которые удаляют нитроны, называют сплайсомами. Они имеют в своём составе несколько малых ядерных РНК (U1—U6), каждая из которых соединена со специфическим белком. Рибонуклеопротеин, содержащий малую ядерную РНК U1 (Ul-малая ядерная РНК), благодаря наличию комплементарной последовательности, присоединяется к участку начала сплайсинга в направлении 3'—5'.
К участку ветви прикрепляется малая ядерная РНК U2, которая затем связывается с U1, в результате чего возникает петля гяРНК. После этого U2 отсекает гяРНК в направлении 3'—5' сразу после последовательности Г—У (см. выше) и соединяет ближний к 5'-конец интрона с участком соединения, образуя так называемое лассо. Конец интрона, находящийся ближе к 3', отсекается сразу после последовательности А—Г, распуская лассо РНК. При этом сплайсома соединяет между собой экзоны.
Иногда в некоторых транскриптах (особенно при производстве антител) обнаруживают альтернативные механизмы сплайсинга, однако ошибки в данном процессе играют важную роль в развитии многих генетических заболеваний. Так, церебральный паралич и задержка умственного развития при синдроме Жильбера обусловлены внедрением Т—А в нормальную последовательность ТАТАА промотора гена УДФ-гликозилтрансферазы. А-аманитин, содержащийся в бледной поганке (Amanita phalloides), блокирует действие РНКазы II.
Антибиотик рифампицин блокирует транскрипцию у бактерий путём связывания с b-субъединицей бактериальной РНК-полимеразы, в то время как актиномицин внедряется между парами оснований Г—С .