Генетика иммуноглобулинов и рецепторов Т-клеток. Особенности экспрессии генов
Большинство примеров, как может измениться экспрессия генов, приходится на изменения транскрипции, альтернативного сплайсинга или посттрансляционной модификации. Активизация или репрессия конкретного гена в данной ткани или в данный период развития обычно связана с изменениями в управлении транскрипцией. Их обеспечивают комбинации специфических факторов транскрипции и других белков, взаимодействующих с управляющими структурами генов в ответ на стимулы внутренней или внешней среды.
В таких случаях сам геном не изменяется, происходит динамическое управление генами, а не изменение их структуры.
Существуют, тем не менее, примеры, когда в результате физической перегруппировки генома или повышенной частоты соматических мутаций изменяются сами гены в клеточных линиях.
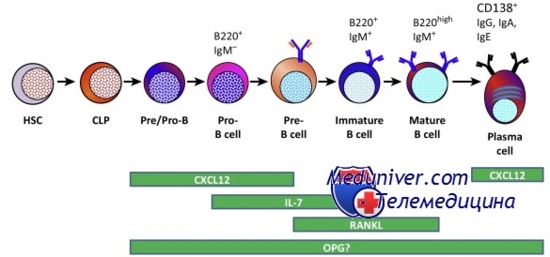
Разнообразие иммуноглобулинов и рецепторов Т-клеток
Антитела, или иммуноглобулины, появляются в ответ на воздействие инородного антигена с целью его распознавания, связывания и устранения. Из-за дефицита иммуноглобулинов возникает множество генетических болезней. С точки зрения генома интересно уникальное свойство иммуноглобулинов — свойство соматической перестройки, благодаря которому в лимфоцитах-предшественниках происходит вырезка-вставка последовательностей ДНК, что и обеспечивает разнообразие антител.
Подсчитано, что каждый человек может генерировать набор около 1011 различных антител, в то время как геном формируется только 6 млрд пар оснований ДНК. Эта кажущееся несоответствие объясняется тем, что изначально антитела кодируются сравнительно небольшим количеством генов, но в ходе развития В-клетки проходят уникальный процесс соматической перестройки и соматических мутаций, которые и обусловливают огромное разнообразие антител.
Молекулы иммуноглобулинов формируются из четырех полипептидных цепей, двух идентичных тяжелых цепей (Н) и двух идентичных легких цепей (L). Каждая Н- и L-цепь белка иммуноглобулина состоит из двух сегментов, постоянного (С) и вариабельного (V). Постоянный участок определяет класс молекулы иммуноглобулина (М, G, А, Е или D), его аминокислотная последовательность сравнительно постоянна среди иммуноглобулинов того же самого класса.
В отличие от этого, аминокислотная последовательность V-участка высоковариабельна у разных антител. V-участки Н- и L-цепей формируют сайт узнавания антигена и определяют специфичность антитела.
Примечательно, что в геноме человека нет полных генов Н- и L-цепей иммуноглобулинов. Как Н-, так и L-цепь кодируется в ДНК половых клеток многочисленными генами, отделенными друг от друга сотнями килобаз. Например, V-участок Н-цепи состоит из трех сегментов, V, D и J. В локусе Н-цепи присутствует более 200 различных генов V-сегмента (хотя некоторые из них, вероятно, псевдогены); далее ниже по хромосоме — приблизительно 30 генов сегмента D и 9 генов сегмента J, сопровождаемые различными генами постоянного участка для каждого типа иммуноглобулинов. В итоге группа генов Н-цепи иммуноглобулина, а также аналогичная группа L-цепи занимают в геноме много миллионов пар оснований.
В ходе дифференцировки клеток, продуцирующих антитела (но не в других клетках!), ДНК в локусах иммуноглобулина перестраивается, создавая функциональные Н- и L-цепи. Для локуса Н-цепи полный ген переменного участка создается разрывом двойной спирали ДНК с последующим соединением свободных концов, приводящим к размещению рядом одного из V-сегментов с одним из D-сегментов, который, в свою очередь, объединяется с одним из J- сегментов, при этом промежуточные участки ДНК удаляются.
Перестроенный сегмент затем транскрибируется, последовательность интронов между вновь сформированным слиянием трех участков экзонов и С-сегментом удаляется, как при обычном сплайсинге. Так возникает зрелая мРНК, готовая для трансляции в специфическую последовательность Н-цепи. Локусы L-цепи подвергаются перед транскрипцией аналогичному процессу перестройки ДНК.
Дополнительное разнообразие антителам придают делеции, вызванные неточным соединением сегментов гена в процессе соматической перестройки. В точках соединения также могут происходить вставки (инсерции), когда включаются нуклеотиды, отсутствовавшие в исходной ДНК (так называемые N-последовательности). Потеря или добавление нескольких нуклеотидов вызывает сдвиг рамки считывания, что приводит к кодированию мутантным геном других аминокислот.
Наконец, когда происходит антигенная стимуляция, В-клетки, производящие антитела с некоторым сходством с конкретным антигеном, начинают размножаться и подвергаться частым точечным мутациям в пределах перестраиваемой последовательности ДНК. Показатель спонтанных мутаций при этом (одна мутация на 103 пары ДНК за деление) поразительно высок, от 100 до 1000 раз больше, чем средний показатель мутаций где-либо еще в геноме.
Спонтанные мутации могут изменять аминокислотную последовательность в переменном (опознающим антиген) участке молекул антитела и тем самым представляют механизм «тонкой настройки» сродства антител к антигену. Разнообразие достигается использованием двух различных цепей (Н и L) ДНК, перегруппированных за счет присоединения различных сегментов первичной ДНК половой клетки (V, D и J), неточного соединения этих сегментов и, наконец, соматических мутаций вариабельного участка. Все перечисленное — крайне важные механизмы расширения потенциального набора специфичности антител.
Механизм соматической перегруппировки используется и другим геном надсемейства генов иммуноглобулинов, геном Т-клеточного рецептора (TCR). TCR — высокоизменчивый трансмембранный гликопротеин, играющий ключевую роль в узнавании антигенов и функции Т-клеток. TCR имеет структурное сходство с молекулой иммуноглобулина; все его цепи имеют как постоянные, так и вариабельные участки, причем последние тоже генерируются набором V, D и J сегментов.
Подобно гену иммуноглобулина, рекомбинация многочисленных элементов первичной ДНК половых клеток, неточности соединения разрывов и возможность различных комбинаций цепей создают широчайшее разнообразие экспрессии гена TCR. Однако, в отличие от иммуноглобулинов, здесь не задействованы соматические мутации.
Соматическая перестройка происходит только в В- и Т-клетках в генах иммуноглобулина и кластере TCR соответственно. Такое поведение уникально для данных семейств генов и клеточных линий; остальная часть генома остается стабильной в ходе развития и дифференцировки.
Аллельное исключение
Соматические перестройки происходят только в одной из двух копий гена иммуноглобулина в В- или TCR в Т-клетке. Это — пример аллельного исключения, когда два аллеля аутосомного локуса обрабатываются по-разному. Основа феномена все еще не до конца понятна.
Хотя большинство аутосомных локусов экспрессируются в обеих копиях гена, есть еще несколько примеров моноаллельной экспрессии. Крайняя форма аллельного исключения обнаружена в семействе генов обонятельных рецепторов, описанном ранее. В этом случае в каждом обонятельном нейроне экспрессируется только один аллель гена, а несколько сотен других копий этого семейства генов остаются в клетке репрессированными.
Для аллельного исключения в локусах иммуноглобулина, TCR и ОР, выбор экспрессирующего аллеля не зависит, от кого из родителей происходит ген, так же как и для генов, подвергающиеся инактивации в Х-хромосоме у женщин, — в разных клетках могут экспрессироваться как материнская, так и отцовская копия. Это отличает аллельное исключение от геномного импринтинга, при котором выбор экспрессирующегося аллеля определяется исключительно родительским происхождением.
Регуляция экспрессии примерно 25 000 генов, закодированных в геноме человека, включает набор сложных взаимозависимостей различных уровней управления, включая точную дозу генов (управляемую механизмами репликации и сегрегации), структуру генов и, наконец, транскрипцию, сплайсинг РНК, устойчивость мРНК, трансляцию, модификацию белка и его распад.
Для некоторых генов колебания в уровне функционального продукта гена, вызванные как унаследованными вариантами в структуре конкретного гена, так и изменениями вследствие негенетических факторов, например диеты или условий среды, играют сравнительно небольшую роль. Для других генов изменения уровня экспрессии могут иметь устрашающие клинические последствия, отражающие значение продуктов гена в конкретных биологических путях. Природа наследуемых изменений в структуре и функции хромосом и генов, их влияние на проявление специфических признаков очень важны для медицинской и молекулярной генетики.