Пикорна-, калици-, астро-, тога- и флавивирусы реплицируются наиболее коротким путем: их (+)РНК геном функционирует непосредственно как мРНК.
Геномы пикорна- и флавивирусов функционируют как единая полицистронная мРНК, транслирующаяся прямо в единый полипротеин, который впоследствии расщепляется с образованием индивидуальных структурных и неструктурных белков. Одним из них является РНК-зависимая РНК-полимераза, которая реплицирует вирусный геном. Вирусная (+)РНК транскрибируется в комплиментарную (—)РНК-копию, которая служит матрицей для синтеза новых цепей (+)РНК (рис. 6), или может использоваться в качестве мРНК-матриц для синтеза новых минус-цепей и геномной РНК вирусного потомства.
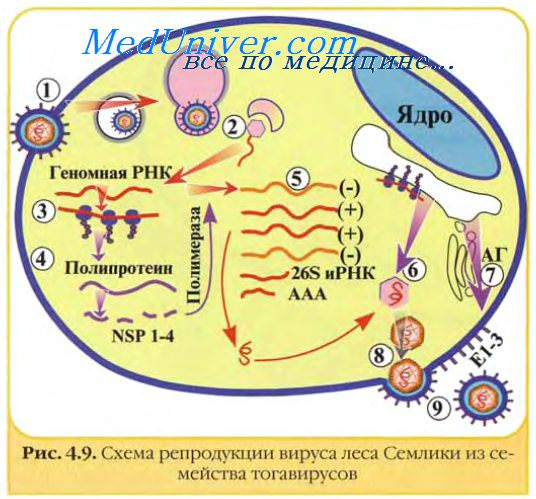
Тога-, корона- и калицивирусы отличаются от пикорнавирусов тем, что на начальном этапе инфекции экспрессируется лишь часть геномной РНК с образованием белков. Последние осуществляют синтез минус-цепи, являющейся матрицей для синтеза различных по размеру классов молекул плюс-РНК. Полипротеины, образующиеся на коротких молекулах мРНК, расщепляются на структурные вирионные белки. Полноразмерные плюс-РНК упаковываются в вирионы.
У тогавирусов транслируется только около 2/3 вирусной РНК (5' -конец); образующийся полипротеин расщепляется на неструктурные белки, которые необходимы для транскрипции и репликации РНК. Вирусная РНК-полимераза синтезирует полноразмерную (—)РНК, на которой затем синтезируются два вида (+)РНК: полноразмерная вирионная РНК, предназначенная для включения в вирионы, и РНК, длина которой равна 1/3, и которая является колинеарной с 3'-концом вирусной РНК и транслируется в полипротеин, который расщепляется на структурные белки. У калицивирусов образуются полигеномные и субгеномные мРНК.
Корона- и артеривирусы демонстрируют необычную стратегию транскрипции: первоначально часть вирионной (+) РНК функционирует как мРНК и транслируется с образованием РНК-полимеразы, которая затем синтезирует полногеномную (—)РНК. На этой (—)РНК транскрибируется гнездо субгеномных мРНК с общими 3'-концами. Транслируются только 5'-концевые последовательности каждого члена этого гнезда транскриптов.
Главным отличием вирусов с позитивным геномом является их способность синтезировать ферменты, ответственные за репликацию вирусного генома. Поэтому РНК, выделенная из таких вирусов, инфекционна. Второе отличие состоит в монолитности вирусного генома. Поэтому первичный продукт трансляции обеих РНК (геномной и мРНК) представляет собой единый белок, который в дальнейшем расщепляется на индивидуальные вирусные белки (в том числе структурные).
Ортомиксо-, бунья- и аренавирусы. Геном представлен (—)РНК. Каждый геномный сегмент транскрибируется отдельно вирионной РНК-транскриптазой и транслируется в один или несколько белков. Эти вирусы характеризуются тем, что их геномная РНК выполняет две матричные функции: в процессе транскрипции и репликации. Транскрипция вирусного генома — первое событие после проникновения вируса в клетку, в результате которого образуются моноцистронные мРНК, кодирующие один белок. Репликацию начинают вновь синтезированные вирусные белки, приводящие к образованию плюс-цепи, которая служит матрицей для синтеза геномной (-)РНК.
Следовательно, плюс-транскрипт, функционирующий в качестве мРНК, отличается от (+)РНК, служащей матрицей для вирусного потомства, хотя и первый, и вторая синтезируются на геномной РНК.
Следует отметить, что S-сегмент РНК аренавирусов и некоторых буньявирусов является двуполярным, т.е. одна часть сегмента имеет (+) полярность, другая (—) полярность. Стратегия репликации двуполярных РНК-вирусов соответствует полярности их геномов и является смешанной, присущей стратегии репликации (+)РНК и (-)РНК вирусов.