ДНК-вирусы. Этапы репликации вирусов с ДНК геномом.
Оспо-, асфаро- и иридовирусы имеют вирионную транскриптазу (ДНК-зависимую РНК-полимеразу) и размножаются в цитоплазме. Их очень большие геномы кодируют многочисленные ферменты, которые делают их репликацию независимой от клеточного ядра. Моноцистронные мРНК транскрибируются непосредственно на вирусной ДНК.
После разрушения наружной оболочки вириона клеточными ферментами на вирусной ДНК с помощью вирионной ДНК-зависимой РНК-полимеразы синтезируются ранние мРНК. Они транслируются в вирусные белки с помощью которых вирионная ДНК полностью освобождается от белка и реплицируется по полуконсервативному механизму. Вначале синтезируются фрагменты ДНК (фрагменты Оказаки), которые затем соединяются с образованием зрелых вирусных ДНК.
Поздние моно- и полицистронные мРНК транскрибируются после начала репликации вирусной ДНК и кодируют синтез структурных белков вириона. Иридовирусы и асфаровирусы, по-видимому, обладают сходной с вирусами оспы стратегией репликации.
Герпес-, адено-, папиллома и полиомавирусы в одном отношении имеют общую стратегию репликации: вирусная ДНК транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. В течение определенного промежутка времени проходит два или более циклов транскрипции различных генных единиц (групп генов под контролем одного промотора). Субгеномные полицистронные РНК-транскрипты подвергаются расщеплению и сплайсингу, превращаясь в моноцистронные мРНК.
У папиллома- и полиомавирусов транскрипционная программа состоит по крайней мере из двух циклов (ранние и поздние мРНК), а у герпес- и аденовирусов по крайней мере из трех (сверхранние, ранние и поздние мРНК). В каждом случае вирионные полипептиды синтезируются на мРНК, появляющейся в последнем цикле транскрипции (поздние мРНК). ДНК аденовирусов, папиллома- и полиомавирусов может встраиваться в геном клетки-хозяина, реплицироваться вместе с ним, передаваться дочерним клеткам и транслироваться с образованием вирусных белков.
Парво- и цирковирусы, обладающие оцДНК, реплицируются в ядре. Эти вирусы используют клеточные ДНК-полимеразы для синтеза двуспиральной ДНК, которая затем транскрибируется в ядре клеточной ДНК-зависимой РНК-полимеразой II. Синтезированные транскрипты подвергаются сплайсингу и превращаются в мРНК.
Вирионная ДНК служит матрицей для синтеза (+) и (-)ДНК. Последние включаются в вирионы в качестве генома.
Различают недефектные (автономные) и дефектные парвовирусы. Первые размножаются самостоятельно, тогда как вторые реплицируются только в присутствии вируса-помощника, которым является любой аденовирус или вирус простого герпеса. В связи с этим дефектные парвовирусы называют аденоассоциированными вирусами (ААВ). В вирионах недефектных парвовирусов содержится односпиральная линейная (+)ДНК. Дефектные парвовирусы являются уникальными в мире вирусов, поскольку комплементарные плюс- и минус-ДНК входят в различные вирионы.
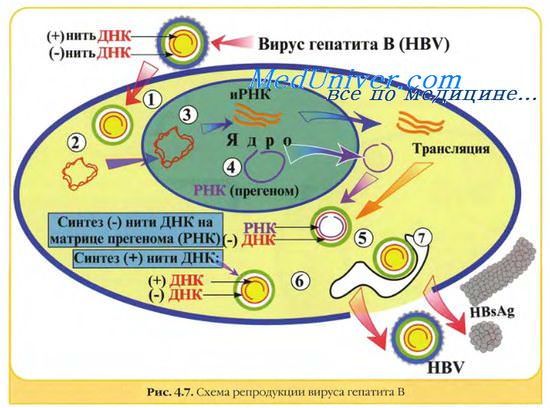
Гепаднавирусы. Частично двуцепочечная ДНК генома гепаднавирусов с помощью вирионной ДНК-полимеразы превращается в суперспирализованную двуцепочечную ДНК. Затем с помощью клеточной РНК-полимеразы II образуются мРНК для вирусных белков, в том числе для обратной транскриптазы, и полноразмерная (+)РНК, которая служит матрицей для обратной вирусной транскрипции с образованием (-)ДНК, на которой в свою очередь синтезируется частично двуцепочечная ДНК. мРНК транскрибируется на двуцепочечной ДНК, стартуя от различных промоторов.