Заселение генитальных складок первичных половых клеток (ППК) происходит одновременно с формированием яичников, с 12-го по 13-й дпк. Мигрировавшие ППК вместе с соматическими эпителиальными клетками, ведущими свое происхождение из целомического эпителия и мезенхимы, формируют яичниковые кортикальные половые тяжи.
Для поддержания жизнеспособности герминативных клеток после 13 дпк необходим фактор транскрипции Zpfl48, который может активировать белок опухолевой супрессии р53 в герминативных клетках данной стадии развития. Приблизительно в это же время ППК дифференцируются в оогонии — уже клетки, прошедшие половую дифференцировку, которые проходят несколько последних раундов митотического деления и формируют популяцию зародышевых клеток с 11-го по 13-й дпк.
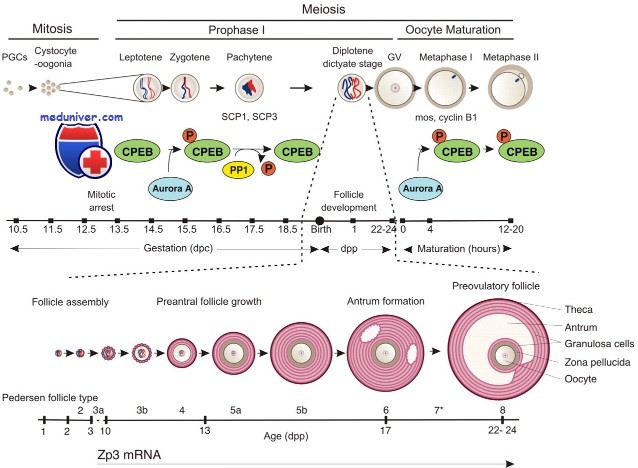
Функции оогониев у млекопитающих не вполне изучены, возможно, потому что их существование весьма кратковременно. Неполный цитокинез в процессе синхронных митотических делений оогониев приводит к образованию кластеров, или цист, соединенных межклеточными мостиками, диаметром 0,5-1,0 мм. Оогонии становятся ооцитами после вхождения в первое деление мейоза с 14-го дпк; к 17-му дпк уже большинство оогониев превращается в ооциты.
В перинатальном периоде герминативные клетки изолируются друг от друга, в результате чего претерпевают растворение, лишь треть ооцитов выживает, и вместе с окружающей однослойной оболочкой гранулезных клеток, формируют примордиальные фолликулы. Таким образом, апоптоз герминативных клеток в перинатальном периоде — один из этапов развития, регулируемых совершенно иначе по сравнению с регуляцией атрезии фолликулов и потери ооцитов, имеющей место у половозрелых животных. Было сделано предположение, что формирование терминального синцития с последующим его распадом необходимо для транспорта митохондрий гибнущих ооцитов в выжившие.
Женская герминативная клетка превращается в ооцит при вхождении в профазу первого мейотического деления. Факторы, сигнализирующие герминативной клетке о начале мейотического деления, до сих пор неизвестны. Даже неизвестно, находятся эти сигналы внутри самой герминативной клетки или в прилегающих соматических клетках. Тем не менее достоверно известно, что к 17-му дпк все оогонии входят в профазу, трансформируясь в ооциты. Репликация ДНК, слияние и рекомбинация хромосом (ключевые признаки полового размножения, обеспечивающие наличие уникального гаплоидного набора в каждой яйцеклетке) происходят именно в профазе.
Профаза I мейоза
Профаза I подразделяется на четыре стадии: лептотену, зиготену, пахитену и диплотену. Репликация ДНК завершается в прелептотену, а в лептотену сестринские хроматиды находят свои гомологичные пары. Образование узелков рекомбинации, облегчающие взаимодействие между гомологичными хромосомами, также начинается в это же время.
На стадии зиготены гомологичные хромосомы объединяются в пары и образуют синапсы. Конъюгация хромосом обеспечивается синаптонемными комплексами, образованными множеством белковых субъединиц, в том числе протеином синаитонемного комплекса 3 (Scp3) в их осевых элементах, Scpl в их центральном элементе и Scp2. Конъюгация завершается в пахитене, иногда сохраняется вплоть до диплотены, когда гомологичные хромосомы соединены главным образом в области хиазм. Кроссинговер и рекомбинация хромосом происходят в течение 4 дней на стадии пахитены перед формированием фолликулов яичника.
В профазе I участвует множество важнейших белков, в том числе белки, проверяющие ДНК на предмет повреждений и исправляющие ошибки в ДНК, в рекомбинации хромосом. На моделях мышей, у которых дефицит этих белков создавался путем нокаута их генов, показаны различные изменения фенотипа — от преждевременного истощения яичников вследствие перинатальной потери ооцитов до дефектов, которые проявлялись позже, в процессе созревания ооцитов.
Эти белки необходимы также и для сперматогенеза, что указывает на общность механизмов профазы I у обоих полов, хотя фенотипическая картина нарушений мейоза у самцов, как правило, более выражена и развивается в более ранние сроки. В этой главе освещено значение указанных белков для оогенеза.