Функции ЛГ и ФСГ. Биохимия и синтез гонадотропных гормонов гипофиза
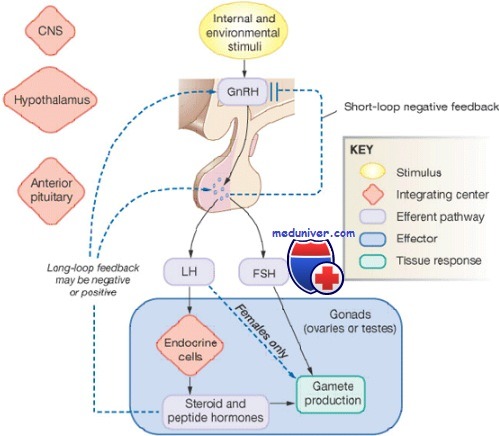
Как было сказано выше, воздействие ГнРГ на гонадотрофы стимулирует продукцию ими гонадотропинов и их высвобождение в импульсном режиме. Кроме того, на высвобождение ЛГ и ФСГ гипофизом влияет секреция эстрогенов, прогестерона и пептидных гормонов яичниками, причем по механизму как отрицательной, так и положительной обратной связи.
Положительное влияние эстрогена и отрицательное — прогестерона зависит от концентрации стероидного гормона и продолжительности воздействия этой концентрации на гонадотрофы. Вместе с тем и ЛГ, и ФСГ необходимы для синтеза стероидов в яичниках, а продукция эстрогена зависит от продолжительности воздействия гонадотропинов и их концентрации.
Во всяком случае дискоординированные сигналы от гипофиза могут привести к урежению овуляции (олигоовуляция) или к ее полному прекращению (ановуляция).
Ранее уже говорилось на нашем сайте МедУнивер, что ЛГ регулирует овариальный стероидогенез. Выброс ЛГ в середине менструального цикла имеет значение в индукции овуляции. Он происходит в результате резкого подъема уровня эстрадиола, производимого преовуляторным фолликулом по механизму положительной обратной связи. Этот выброс стимулирует возобновление мейоза и завершение редукционного деления ооцита с образованием первого полярного тела.
В ответ на выброс ЛГ повышается уровень протеолитических ферментов и простагландинов, способствующих выходу яйцеклетки из яичника. В конечном счете продолжающаяся секреция ЛГ после овуляции превращает оставшиеся фолликулярные клетки в желтое тело и стимулирует продукцию им прогестерона, интенсифицируя превращение холестерина в прегненолон.
ФСГ необходим для созревания и роста фолликулов; он регулирует синтез овариальных эстрогенов, связываясь со своими рецепторами на поверхности гранулезных клеток. Это приводит к повышению уровня циклического аденозинмонофосфата (цАМФ) и индукции ароматазы, которая превращает андростендион соседних текальных клеток в эстрон. ФСГ также запускает экспрессию 17PHSDH 1-го типа, которая конвертирует эстрон в эстрадиол.
Повышенная секреция эстрадиола способствует дальнейшей пролиферации гранулезных клеток и росту фолликулов, а также увеличению числа эстрогеновых рецепторов. В зрелом фолликуле ФСГ и эстрадиол повышают экспрессию рецепторов к ЛГ на гранулезных клетках, делая их более чувствительными к ЛГ и усиливая секрецию прогестерона. Прогестерон затем увеличивает высвобождение ФСГ в середине цикла.
Биохимия и биосинтез ЛГ и ФСГ - гонадотропных гормонов гипофиза
Передняя доля гипофиза продуцирует три гликопротеиновых гормона: ЛГ, ФСГ и тиреотропинстимулирующий гормон (ТСГ); все они имеют сходную биохимическую структуру. Каждый гормон состоит из гетеродимера, содержащего две нековалентно связанные протеиновые субъединицы а и b. Эти субъединицы богаты цистеином и содержат множество дисульфидных связей.
На обеих субъединицах имеется множество углеводородных компонентов, которые играют важную роль в метаболизме и биологической активности гормонов. а-Субъединица является общей для всех трех гормонов, в то время как b-субъединица у всех разная.
а-Субъединица гонадотропных гормонов гипофиза
Ген а-субъединицы человека локализован на коротком плече хромосомы 6 (локус 6р21.1-23) и состоит из 4 экзонов. Первый экзон некодирующий. Ген кодирует полипептид с молекулярной массой 14 кДа, состоящий из 24-аминокислотного сигнального пептида и 92-аминокислотной зрелой а-субъединицы с 10 цистеиновыми остатками и двумя N-связанными олигосахаридными группами. Цистеиновые остатки участвуют в образовании внутренних дисульфидных связей а-субъединиц больше, чем р-субъединиц, и несвязанные, свободные а-субъединицы присутствуют в крови и гипофизе.
Какой-либо известной биологической активностью они не обладают. Иными словами, биологически активен лишь а/b-гетеродимер.
b-Субъединица ЛГ и ФСГ - гонадотропных гормонов гипофиза
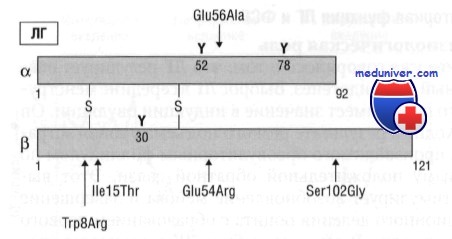
b-Субъединицы ЛГ и ФСГ кодируются разными генами, расположенными на разных хромосомах. Ген, кодирующий ЛГb-субъединицу, состоит из трех экзонов и входит в комплексный генный кластер хромосомы 19 человека (локус 19ql3.3). Этот кластер также включает шесть генов хорионического гонадотропи - на b (ХГр), предположительно ведущих свое происхождение от единого предшественника — гена ЛГb-субъединицы, который подвергся дупликации.
Оба белка, ЛГb и ХГb, структурно и функционально сходны. Они приблизительно на 80% гомологичны по аминокислотной последовательности, оба содержат 20-аминокислотную сигнальную последовательность. Зрелые ЛГb- и ХГb-субъединицы состоят соответственно из 121 и 145 аминокислот. Основное различие между ЛГb-и ХГb-протеинами — наличие 24-аминокислотного С-терминального пептида у ХГb, обильно гликозилированного, с четырьмя кислородсвязанными углеводородными компонентами.
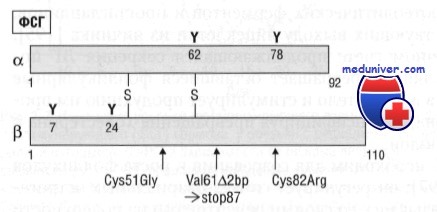
Ген ФСГb-субъединицы расположен на коротком плече хромосомы 11 (локус 11р13) и, как и ген ЛГ, состоит из трех экзонов. Молекулярная масса ФСГ составляет 33 кДа; он состоит из 18-аминокислотного сигнального пептида и зрелого 111-аминокислотного ФСГр-протеина. Как и a-субъединицы, ЛГb- и ФСГb-субъединицы содержат по 10 цистеиновых остатков для образования дисульфидных связей. В отличие от ФСГb-и ХГb-субъединиц, ЛГb-субъединица не имеет терминальной сиаловой кислоты на углеводородной боковой цепи. Из-за этого продолжительность метаболического клиренса ЛГ меньше, чем ФСГ и чХГb-субъединиц.
Самым большим содержанием сиаловой кислоты характеризуется чХГb-субъединица, что выражается в максимальном периоде полужизни чХГ. Дегликозилирование гонадотропинов не влияет на связывание с рецепторами, но препятствует передаче сигнала.