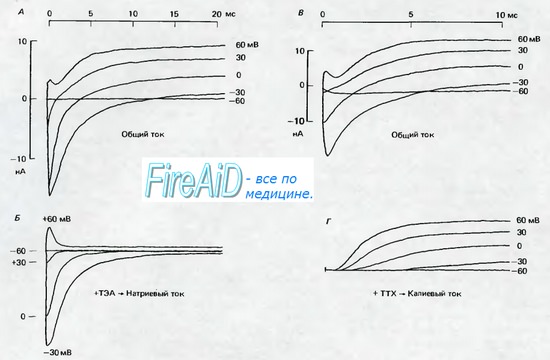
Рис. 2.6. А и В Мембранные токи в миелинизированных аксонах лягушки (перехваты Ранвье; 11 13 С) после ступенчатых сдвигов мембранного потенциала. Мембранный потенциал поддерживался с помощью фиксации потенциала на уровне потенциала покоя, равного —95 мВ; в момент времени 0 мс мембранный потенциал скачком поднимали до значений, которые указаны справа около записей тока, от —60 до +60 мВ. Сопровождающие скачок потенциала кратковременные емкостные токи вычитались, поэтому регистрируемые токи являются ионными токами. При —60 мВ скачок потенциала остается подпороговым и не вызывает изменений тока. По мере увеличения скачков потенциала сначала возникают отрицательные токи, которые с увеличением потенциала становятся положительными. Б. То же, что на рис. А, но на фоне блокады калиевых токов ТЭА (6 мМ), в результате чего токи почти полностью обеспечиваются ионами Na+. Полярность Na-токов меняется с отрицательной на положительную между значениями +30 и +60 мВ; по мере увеличения деполяризации продолжительность Na - токов уменьшается. Г. То же, что на рис. В. но на фоне блокады натриевых токов тетродотоксином (0,3 мкМ), так что записи соответствуют калиевым токам. При деполяризации К+-токи нарастают медленнее, чем Na - токи, и продолжаются в течение всего периода деполяризации (по [3] с изменениями)
Из рис. 2.6, Б видно, что натриевый (Na+)-ток в нерве лягушки начинает убывать примерно через 0,5 мс, несмотря на продолжающуюся деполяризацию. У гомойо-термных животных, имеющих более высокую температуру тела, этот промежуток времени еще короче. Характерная для Na-тока инактивация становится более быстрой по мере увеличения деполяризации, так что ток раньше возвращается к нулю. Однако этот процесс не означает восстановления состояния покоя; если мембрана на короткое время реполяризуется и снова деполяризуется после полной инактивации, в ней практически невозможно снова вызвать натриевый (Na+)-ток. При таком состоянии мембраны натриевая система не может быть активирована.
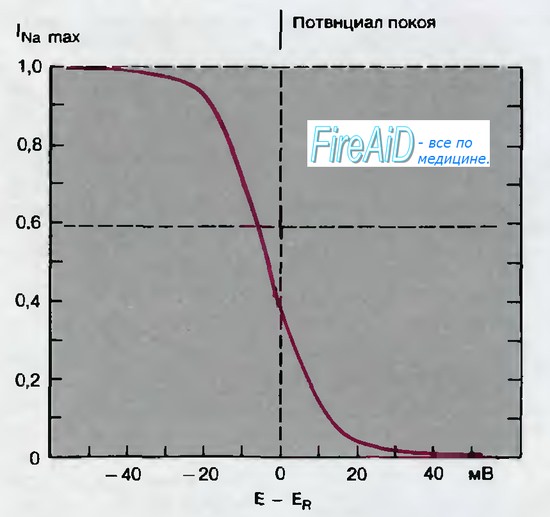
Рис. 2.8. Потенциал зависимая инактивация натриевой (Na)-системы. По оси абсцисс отложены величины отклонения мембранного потенциала от потенциала покоя ( — 60 мВ). От каждого из этих исходных значений потенциала мембрану деполяризовали до —16 мВ и по оси ординат откладывали отношения возникающих максимальных натриевых (Na+) -токов (lNamax) к величине lNamax, соответствующей полной активации натриевой (Na)-системы (по [15] с изменениями)
Даже после того как на некоторое время восстановится потенциал покоя, натриевый (Na+) - ток можно активировать лишь частично. Только в случае предварительной гиперполяризации аксонной мембраны на 20-40 мВ последующая деполяризация от этого уровня потенциала способна вызвать максимальный натриевый (Na+)-ток, I(Namax) (рис. 2.8). Если мембранный потенциал на период 10 мс или дольше сместить на 20 мВ от потенциала покоя к более положительному значению, то деполяризация от этого исходного уровня приведет к возникновению только минимального натриевый (Na+)-тока. Следовательно, длительная деполяризация может предотвратить возбуждение; клетки, потенциал которых положительнее уровня от —60, до —50 мВ, утрачивают возбудимость [3, 23]. Продолжительная деполяризация может развиваться в результате метаболических нарушений, например при кислородной недостаточности, а также под влиянием фармакологических препаратов: такая деполяризация способна блокировать генерирование процесса возбуждения.