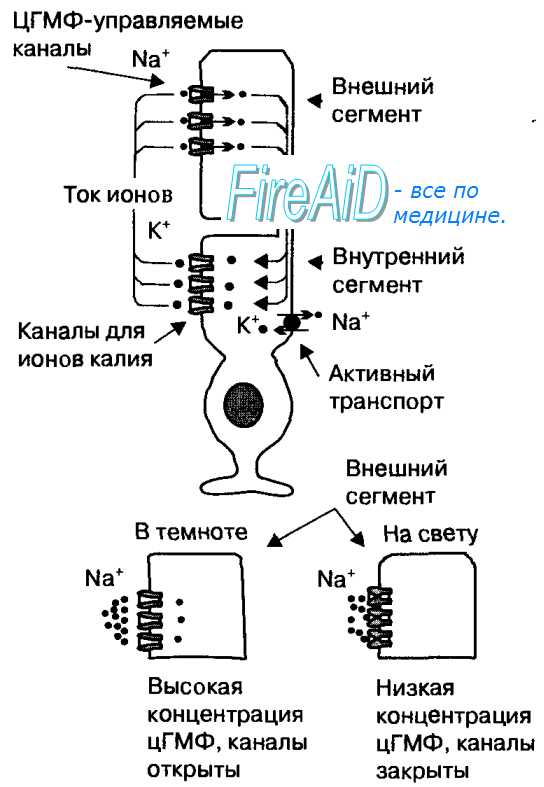
Рецепторный потенциал палочек и колбочек. Ток ионов через мембрану фоторецептора в темноте и на свету.
Специфической особенностью фоторецепторов является темновой ток катионов через открытые мембранные каналы внешних сегментов (рис. 17.8). Эти каналы открываются при высокой концентрации циклического гуанозинмонофосфата, который является вторичным посредником рецепторного белка (зрительного пигмента). Темновой ток катионов деполяризует мембрану фоторецептора до приблизительно —40 мВ, что приводит к выделению медиатора в его синаптическом окончании. Активированные поглощением света молекулы зрительного пигмента стимулируют активность фосфодиэстеразы — фермента, расщепляющего цГМФ, поэтому при действии света на фоторецепторы в них уменьшается концентрация цГМФ. В результате управляемые этим посредником катионные каналы закрываются, и ток катионов в клетку прекращается.
Рис. 17.8. Ток ионов через мембрану фоторецептора в темноте и на свету.
Ионы натрия поступают в фоторецепторы через катионные каналы внешнего сегмента, которые открываются при высокой концентрации цГМФ в темноте (темновой ток). Поступление в клетку ионов натрия вызывает деполяризацию мембраны фоторецептора до —40 мВ. На свету концентрация цГМФ уменьшается, что приводит к закрытию катионных каналов. Непрекращающийся выход ионов калия ведет к гиперполяризации мембраны, которая представляет собой рецепторный потенциал.
Вследствие непрерывного выхода ионов калия из клеток, мембрана фоторецепторов гиперполяризуется приблизительно до —70 мВ, эта гиперполяризация мембраны является рецепторным потенциалом. При возникновении рецепторного потенциала прекращается выделение глутамата в синаптических окончаниях фоторецептора.
Фоторецепторы образуют синапсы с биполярными клетками двух типов, различающихся по способу управления хемозависи-мыми натриевыми каналами в синапсах. Действие глутамата приводит к открытию каналов для ионов натрия и деполяризации мембраны одних биполярных клеток и к закрытию натриевых каналов и гиперполяризации биполярных клеток другого типа. Наличие двух типов биполярных клеток необходимо для формирования антагонизма между центром и периферией рецептивных полей ганглиозных клеток.