Активный транспорт, натриевый насос. В предыдущем разделе описаны пассивная диффузия ионов и образующийся вследствие этого мембранный потенциал при заданных внутри- и внеклеточных концентрациях ионов. Однако в результате этого процесса концентрация ионов внутри клетки стабилизируется не автоматически, поскольку мембранный потенциал несколько более электроотрицателен, чем ЕK, и намного - по сравнению с ENa (около +60 мВ). Благодаря диффузии внутриклеточные концентрации ионов, по крайней мере калия и натрия, должны уравниваться с внеклеточными.
Стабильность градиента ионов достигается посредством активного транспорта: мембранные белки переносят ионы через мембрану против электрического и (или) концентрационного градиентов, потребляя для этого метаболическую энергию. Наиболее важный процесс активного транспорта - это работа Na/K-насоса, существующего практически во всех клетках; насос выкачивает ионы натрия из клетки, одновременно накачивая ионы калия внутрь клетки. Таким образом обеспечивается низкая внутриклеточная концентрация ионов натрия и высокая-калия (табл. 1.1). Градиент концентрации ионов натрия на мембране имеет специфические функции, связанные с передачей информации в виде электрических импульсов, а также с поддержанием других активных транспортных механизмов и регулирования объема клетки (см. ниже). Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой, расходуется на Na/К-насос, а в некоторых наиболее активных клетках на его работу расходуется до 70% энергии [1, 11].
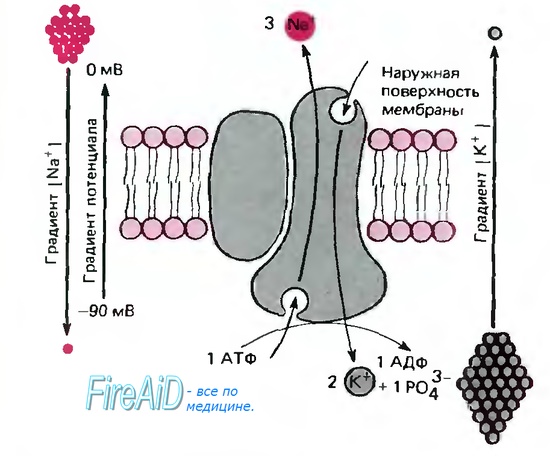
Рис 1.6. Схема Na/K-насоса АТФазы (погруженной в липидный бислой плазматической мембраны), которая за один цикл выносит из клетки три иона Na+ против градиентов потенциала и концентрации и приносит в клетку два иона К+. В ходе этого процесса одна молекула АТФ расщепляется на АДФ и фосфат. На схеме АТФаза показана как димер, состоящий из большой (функциональной) и малой субъединиц; в мембране она существует как тетрамер, образованный двумя большими и двумя малыми субъединицами
Na/K-транспортный белок представляет собой АТФазу. На внутренней поверхности мембраны она расщепляет АТФ на АДФ и фосфат (рис. 1.6). На транспортировку трех ионов натрия из клетки и одновременно двух ионов калия в клетку используется энергия одной молекулы АТФ, т. е. суммарно за один цикл из клетки удаляется один положительный заряд. Таким образом, Na/К-насос является электрогенным (создает электрический ток через мембрану), что приводит к увеличению электроотрицательности мембранного потенциала приблизительно на 10 мВ. Транспортный белок выполняет эту операцию с высокой скоростью: от 150 до 600 ионов натрия в секунду. Аминокислотная последовательность транспортного белка известна, однако еще не ясен механизм этого сложного обменного транспорта. Данный процесс описывают с использованием энергетических профилей переноса белками ионов натрия или калия (рис. 1.5,-6). По характеру изменения этих профилей, связанных с постоянными изменениями конформации транспортного белка (процесс, требующий затраты энергии), можно судить о стехиометрии обмена: два иона калия обмениваются на три иона натрия.
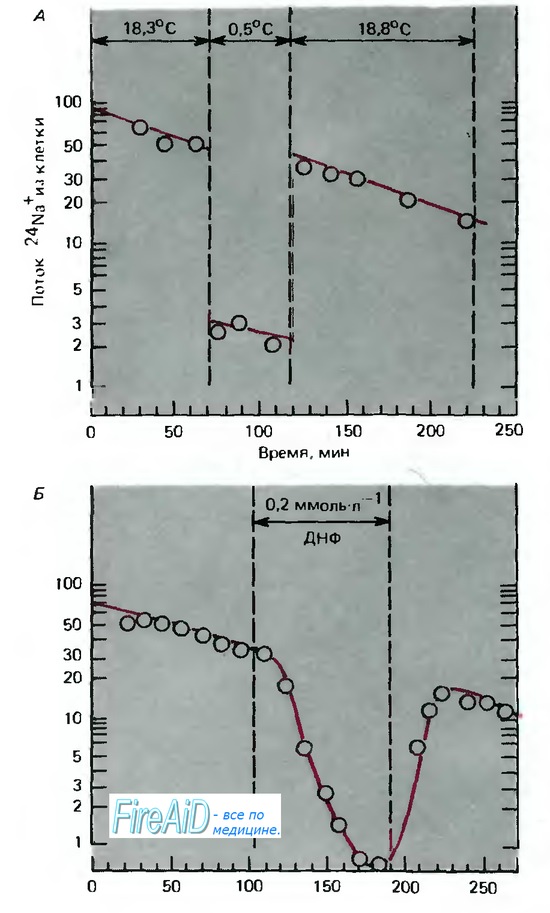
Na/К-насос, как и изолированная Na+/К-зависимая мембранная АТФаза, специфически ингиби-руется сердечным гликозидом уабаином (строфантином). Поскольку работа Na/K-насоса представляет собой многоступенчатую химическую реакцию, она, подобно всем химическим реакциям, в значительной степени зависит от температуры, что продемонстрировано на рис. 1.7. Здесь поток ионов натрия из мышечных клеток показан относительно времени; практически это эквивалентно потоку ионов натрия, опосредованному работой Na/K-насоса, потому что пассивный поток ионов натрия против градиентов концентрации и потенциала крайне мал. Если препарат охладить примерно на 18 С, то поток ионов натрия из клетки быстро уменьшится в 15 раз, а сразу после нагревания восстановится до исходного уровня. Такое уменьшение потока ионов натрия из клетки в несколько раз больше, чем то, которое бы соответствовало температурной зависимости процесса диффузии или простой химической реакции. Сходный эффект наблюдается, когда запас метаболической энергии истощается в результате отравления динитрофенолом (ДНФ) (рис. 1.7, Б). Следовательно, поток ионов натрия из клетки обеспечивается энергозависимой реакцией - активным насосом. Другой характеристикой насоса наряду со значительной температурной и энергетической зависимостью является наличие уровня насыщения (как и у всех других химических реакций); это означает, что скорость работы насоса не может возрастать бесконечно при повышении концентрации транспортируемых ионов (рис. 1.8). В отличие от этого поток пассивно диффундирующего вещества растет пропорционально разности концентраций в соответствии с законом диффузии (уравнения 1 и 2).
Рис. 1.7. А, Б Активный транспорт Na+. Ось ординат поток радиоактивного 24Na+ из клетки (имг.-мин-1). Ось абсцисс: время с начала эксперимента. А. Клетка охлаждена с 18,3c до 0,5 С; поток Na+ из клетки в этот период заторможен.
Б. Подавление потока Na+ из клетки динитрофенолом (ДНФ) в концентрации 0-2 ммоль/л (по [13] с изменениями)
Помимо Na/K-насоса плазматическая мембрана содержит по крайней мере еще один насос—кальциевый; это насос откачивает ионы кальция (Са2+) из клетки и участвует в поддержании их внутриклеточной концентрации на крайне низком уровне (табл. 1.1). Кальциевый насос присутствует с очень высокой плотностью в саркоплазматическом рети-кулуме мышечных клеток, которые накапливают ионы кальция в результате расщепления молекул АТФ.