Клеточная стенка бактерий. Функции клеточной стенки. Строение клеточной стенки бактерии. Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
Основные функции клеточной стенки следующие.
• Клеточная стенка защищает бактерии от внешних воздействий, придаёт им характерную форму, поддерживает постоянство внутренней среды и участвует в делении.
• Через клеточную стенку бактерий осуществляется транспорт питательных веществ и выделение метаболитов,
• На поверхности клеточной стенки располагаются рецепторы для бактериофагов, бактериоци-нов и различных химических веществ.
• Структура и состав элементов клеточной стенки определяет антигенную характеристику бактерий (по структуре О- и Vi-Аг).
• Клеточная стенка способна по-разному воспринимать красители; на этом основаны тинкториальные свойства бактерий.
• Нарушение синтеза компонентов клеточной стенки приводит к гибели бактерии или образованию 1-форм.
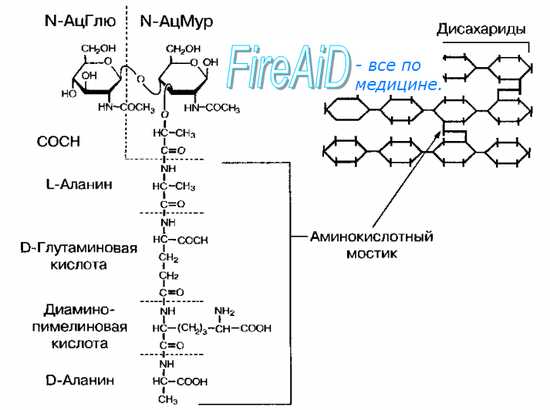
Рис. 4-3. Структура пептидогликана (муреина) Escherichia coll. Гетерополимерные цепочки, состоящие из чередующихся остатков N-ацитилглюкозамина (N-АцГлю) и N-ацетилмурамовой кислоты (N-АцМур), связаны между собой аминокислотными мостиками. Слева— детальное строение фрагмента пептидогликана. Справа — схематическое изображение структуры однослойного муреи-нового мешка.
Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
Опорный каркас клеточной стенки бактерий — пептидогликан (муреин) — гетерополимер, состоящий из повторяющихся дисахаридных групп, соединённых поперечными и боковыми цепочками (рис. 4-3). Остов молекулы пептидогликана — дисахарид. Его образуют N-ацетилглюкозамин и N-ацетилмурамовая кислота, соединённые через р-гликозидные связи. К молекуле N-ацетилмурамовой кислоты присоединяются олигопептиды, образующие боковые цепочки.
Связывание фрагментов пептидогликана заключается в образовании пептидной связи между терминальным остатком аминокислотного мостика (D-аланином) с предпоследним остатком примыкающего аминокислотного мостика (L-лизином или диаминопимелиновой кислотой в зависимости от вида бактерии). Боковые мостики образуют четыре аминокислоты, поперечные (вертикально связывающие слои пептидогликана) — пять аминокислот. В состав боковых мостиков также входят уникальные аминокислоты, присутствующие только у прокариотов, например диаминопимелиновая кислота (у большинства грамотрицательных бактерий) и D-изомеры глутаминовой кислоты и аланина. Эти соединения — «ахиллесова пята» бактерий, так как препараты, ингибирующие синтез компонентов клеточной стенки, обычно безвредны для растений и животных.
Пептидогликан лабилен к действию различных агентов. В частности, лизоцим гидролизует пептидогликан, расщепляя гликозидные связи между N-ацетилглюкозамином и N-ацетилмурамовой кислотой, то есть лизоцим действует как N-ацетилмурамидаза. Пептидазы расщепляют межпептидные связи, амидазы — связи между N-ацетилмурамовой кислотой и боковым пептидом (L-аланином). Лизостафин разрушает поперечные мостики пептидогликанов стафилококков. В-Лактамные антибиотики нарушают связывание боковых мостиков.
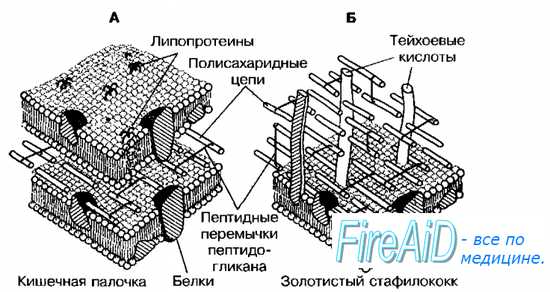
Рис. 4-4. Клеточная стенка грамотрицательных (А) и грамположительных (Б) бактерий.
Гетерополимерные пептидогликановые цепочки, соединённые пептидными связями, образуют гигантскую мешковидную макромолекулу («муреиновый мешок»), покрывающую всё тело бактерии. На поверхности «муреинового мешка» и в его толще располагаются различные вещества, характер и содержание которых лежит в основе разделения бактерий на грамположительные и грамотрицателъные (рис. 4-4).