Слово «хромосома» означает «окрашенное тело» (окрашивается более эффективно, чем остальные компоненты клетки). Каждая хромосома состоит из очень длинной молекулы ДНК, связанной с РНК и белками, образующей вещество, известное как хроматин. Во время интерфазы клеточного цикла (когда клетка не делится) хромосомы распределены по всему ядру, в то время как при митозе и мейозе они уплотняются, располагаясь ближе друг к другу.
ДНК содержится именно в хромосомах, вероятно, потому, что это облегчает разделение полного набора генов между дочерними клетками на стадии митоза, а также её упаковку в головки сперматозоидов после мейоза.
Способность к окрашиванию используют в диагностических целях для визуального исследования хромосом, их идентификации и выявления патологий. Различают слабоокрашенные части (эухроматин) и области с более выраженной окраской (гетерохроматин).
Генетическая информация (геном) заключена в молекуле ДНК и кодирована определённой последовательностью азотистых оснований в ДНК. Основная часть этой информации находится в составе хромосом в ядре клетки, однако небольшое количество ДНК в виде свободных цепочек локализовано в митохондриях цитоплазмы. Ядра присутствуют практически во всех клетках организма человека, за исключением эритроцитов и клеток хрусталика.
Обычно в каждом ядре содержится двухметровая цепь ДНК, разделённая между 23 парами хромосом (примерно по 4 см на хромосому). Однако непосредственно перед делением она уплотняется до 5 um (0,005 мм) посредством сложных процессов спирализации и уплотнения.
Структура хроматина хромосом
В каждой хромосоме цепь ДНК дважды обмотана вокруг октамерного комплекса, состоящего из восьми «белков хранения» (гистонов), формирующих нуклеосомы, при этом образованная структура напоминает «бусы на нити». Кора (сердцевина) нуклеосомы состоит из двух молекул каждого из четырех гистонов— Н2А, Н2В, НЗ и Н4. Гистоны заряжены положительно, а потому могут образовывать ионные связи с отрицательно заряженными фосфатными группами ДНК.
Последовательность аминокислот в гистонах у всех видов совпадает практически на 100%, указывая на огромную значимость данных элементов в поддержании структуры и функций хроматина. Каждая нуклеосома вмещает порядка 200 пар оснований молекулы ДНК и укорачивает длину цепи ДНК на одну десятую.
Затем подобная бусам нить спирализуется в соленоид, или зигзагообразную спираль (фибрилла длиной 30 нм), состоящую из 5—6 нуклеосом. При этом её структура поддерживается одной молекулой гистона HI на каждую нуклео-сому. Коэффициент упаковки при образовании соленоида примерно равен 5, что в свою очередь даёт общий коэффициент упаковки 50. Существует предположение, что в таком состоянии эухроматин находится во время интерфазы в местах, где гены не экспрессируются.
Во время митоза и мейоза хромосомы ещё больше уплотняются, при этом коэффициент упаковки по отношению к предыдущим уровням составляет 100, а общий коэффициент может достигать 5000. Считают, что фибриллы хроматина складываются в вытянутые петли, в основании которых расположен скаффолд (остов) из негистоновых хромосомных белков, которые прикрепляются к определённым последовательностям оснований, рассредоточенным вдоль молекулы ДНК. Сжатие этих негистоновых хромосомных белков предположительно обусловливает уплотнение хромосом.
Один из ключевых белков скаффолда — топоизомераза II, фермент, который расщепляет двойные цепи ДНК, переносит их через разрыв и вновь соединяет, что в свою очередь необходимо для релаксации суперспиралей ДНК во время репликации или транскрипции. Топоизомераза II прикрепляется к участкам связывания со скаффолдом, которые богаты А- и Т-основаниями (более 65% оснований составляют А и Т, см. главу 4). Считают, что каждая петля выступает в роли независимого функционального домена во время репликации или транскрипции ДНК.
На следующем этапе петли фибрилл хроматина спирализуются, в результате чего возникает наиболее уплотнённый гетерохроматин, входящий в состав хромосомы во время деления клеток.
Дифференциальное окрашивание хромосом
Наиболее уплотнённые части хромосом хорошо окрашиваются красителем Гимзы (G-окраска). При этом отчётливо видны сильно уплотнённые маленькие петли (G-полосы), так как участки связывания со скаффолдом тесно прилегают друг к другу. Их репликация происходит во время S-периода, они не активны во время транскрипции. Полосы, которые плохо окрашиваются раствором Гимзы, или R-полосы, состоят из более свободных петель, богатых Г- и Ц-основаниями, они отличаются высокой активностью во время транскрипции. В основе идентификации хромосом лежат различия в их окраске.
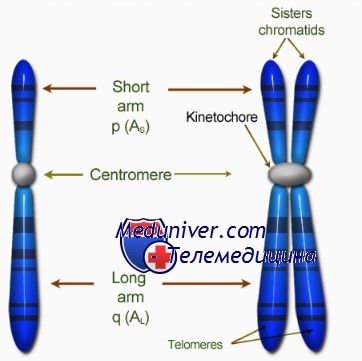
Центромера хромосом
Видимые на ранних стадиях митоза хромосомы состоят из двух абсолютно одинаковых структур, называемых сестринскими хроматидами, которые соединяет первичная перетяжка. Она представляет собой недуплицированный участок ДНК, или центромеру, которая дуплицируется в начале анафазы митоза.
Кинетохор — органелла, которая локализована на каждой стороне каждой центромеры в период ранней профазы митоза и облегчает полимеризацию димеров тубулина, необходимую для образования микротрубочек митотического веретена (веретена деления).
Тепомера хромосом
Теломерой называют специализированный участок конца хромосомы. К ней прикрепляются теломерспецифические белки, образующие «шапочку» (cap) для защиты конца хромосомы.
Теломеры предположительно препятствуют патологическому слиянию концов хромосом конец в конец, обеспечивают полноту репликации, участвуют в образовании пар хромосом во время мейоза и помогают восстанавливать внутреннюю структуру ядра на стадии интерфазы путём присоединения хромосом к ядерной мембране.
Эухроматин и гетерохроматин хромосом
Эухроматин находится в уплотнённом состоянии во время деления клеток и распаковывается на стадии интерфазы. Он обусловливает бледное окрашивание R-полос в уплотнённых хромосомах и содержит большинство структурных генов.
Гетерохроматин очень сильно уплотнён во время деления клетки и остаётся в таком состоянии даже на стадии интерфазы. Он локализован в основном на периферии ядра и около ядрышка и не активен во время транскрипции. Конструктивный (структурный) гетерохроматин одинаков во всех клетках организма, в то время как факультативный гетерохроматин различен в зависимости от типа генов, экспрессируемых в дифференцированных клетках определённых тканей.