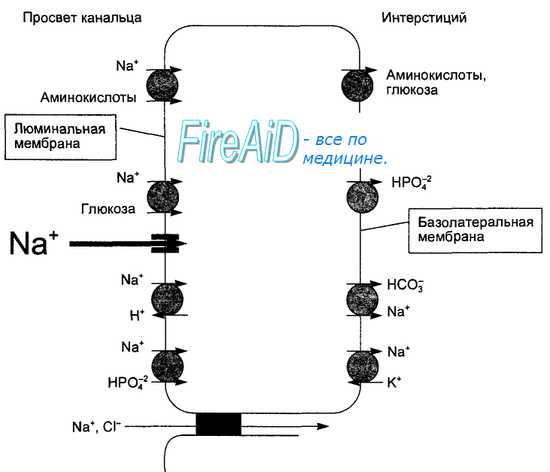
Реабсорбция ионов натрия в проксимальном отделе осуществляется несколькими механизмами активного и пассивного транспорта (рис. 14.9). Во-первых, реабсорбция натрия осуществляется первично активным транспортом. Ионы натрия входит в клетки эпителия через апикальную мембрану пассивно через натриевые каналы по концентрационному градиенту, его выведение через базолатеральные мембраны эпителиальных клеток происходит активно с помощью натрий-калиевых насосов, использующих энергию АТФ. Именно деятельность этих насосов обеспечивает градиент концентрации ионов натрия между внутриканальцевой и внутриклеточной средами. Во-вторых, на апикальной мембране имеется электронейтральный переносчик, обеспечивающий активный обмен Na+ и Н+, при этом ион натрия поступает в клетку в обмен на удаляемый из клетки Н-ион. Такой механизм транспорта носит название антипорта.
Этот переносчик обеспечивает и всасывание бикарбонатного аниона. Профильтровавшийся бикарбонатный анион вместе с Н-ионом образуют угольную кислоту: HCO3 + Н+ = Н2С03. Располагающаяся на щеточной каемке эпителия канальца карбоангидраза катализирует разложение в канальцевой жидкости угольной кислоты: Н2С03 о Н20 + С02, после чего С02 диффундирует в клетку по градиенту концентрации. В клетке под влинием цитоплазменной карбоангидразы протекает обратная реакция: С02 + Н20=Н2С03, угольная кислота диссоциирует: Н2С03 о Н+ + HCO3. Бикарбонатный анион (НСОз) пассивно переносится в перитубулярную жидкость по электрохимическому градиенту, создаваемому активным переносом натрия через ба-золатеральную мембрану, а Н-ион через апикальную мембрану с помощью антипорта Na+-H+ выводится в просвет канальца. Таким образом, сопровождающим всасывающийся ион натрия в начальных отделах проксимального канальца анионом является бикарбонат. Анионы хлора всасываются в начальных отделах плохо из-за низкой проницаемости стенки. Объем мочи в канальце уменьшается из-за пассивной реабсорбции воды, и концентрация хлоридов в его содержимом растет. В конечных участках проксимальных канальцев межклеточные контакты уже проницаемы для хлоридов (концентрация которых повысилась) и они пассивно по градиенту концентрации всасываются из мочи путем парацеллюлярной диффузии, создавая электрохимический градиент для натрия. В-третьих, ион натрия реабсорбируется пассивно, по электрохимическому градиенту, вслед за анионом хлора. Такой пассивный транспорт одного иона (натрия) вместе с пассивным транспортом другого (хлорида) носит название котранспорта. В-четвертых, на апикальной мембране расположены переносчики-котранспортеры натрия и органических веществ (глюкозы, аминокислот), натрия и фосфата или сульфата.
Рис. 14.9. Основные системы транспорта натрия в проксимальном канальце нефрона.
Жирной стрелкой выделено поступление натрия в клетку через натриевый канал по концентрационному градиенту (пассивный транпорт путем простой диффузии). На люминальной мембране черными кружками обозначены переносчики, осуществляющие вторично активный котранспорт путем облегченной диффузии (Na+ и глюкоза, Na+ и аминокислоты), либо пассивный котранспорт (Na+ и фосфат), либо антипорт (Na+ и Н+). На базолатеральной мембране расположены насосы, обеспечивающие активный транспорт натрия из клетки (Na+-K+-Hacoc). Черный квадрат — основное вещество плотного межклеточного соединения, при деполимеризации которого возможен пассивный межклеточный транспорт Na.
Проксимальная реабсорбция глюкозы и аминокислот осуществляется с помощью специальных переносчиков щеточной каемки апикальной мембраны эпителиальных клеток. Эти переносчики транспортируют глюкозу или аминокислоту, только если одновременно связывают и переносят натрий. Пассивное перемещение натрия по градиенту концентрации внутрь клеток ведет к транспорту через мембрану и переносчика с глюкозой или аминокислотой. Для реализации этого процесса необходима низкая концентрация натрия в эпителиальной клетке, создающая градиент концентрации между внешней и внутриклеточной средой, что обеспечивается энергозависимой работой натрий-калиевого насоса базальной мембраны. Поскольку перенос глюкозы или аминокислоты связан с натрием, а его транспорт определяется активным удалением натрия из клетки, такой вид транспорта называют вторично активным, или симпортом, т. е. совместным пассивным транспортом одного вещества (глюкоза) из-за активного транспорта другого (натрия) с помощью одного переносчика.
Ввиду того что для реабсорбции глюкозы необходимо связывание каждой ее молекулы с молекулой переносчика, при избытке глюкозы в первичной моче может произойти полная загрузка всех молекул переносчиков и глюкоза уже не сможет всасываться в кровь. Эта ситуация характеризуется понятием «максимальный канальцевый транспорт вещества» (Тм глюкозы), которое отражает максимальную загрузку канальцевых переносчиков при определенной концентрации вещества в первичной моче и, соответственно, в крови. Эта величина составляет от 303 мг/мин у женщин до 375 мг/мин у мужчин. Величине максимального канальцевого транспорта соответствует понятие «почечный порог выведения».
Почечным порогом выведения называют ту концентрацию вещества в крови и, соответственно, в первичной моче, при которой оно уже не может быть полностью реабсорбировано в канальцах и появляется в конечной моче. Такие вещества, для которых может быть найден порог выведения, т. е. реабсорбирующиеся при низких концентрациях в крови полностью, а при повышенных концентрациях — не полностью, носят название пороговых. Примером является глюкоза, которая полностью всасывается из первичной мочи при концентрациях в плазме крови ниже 10 ммоль/л, но появляется в конечной моче, т. е. полностью не реабсорбируется, при содержании ее в плазме крови выше 10 ммоль/л. Следовательно, для глюкозы порог выведения составляет 10 ммоль/л.
Вещества, которые вообще не реабсорбируются в канальцах (инулин, маннитол) или мало реабсорбируются и выделяются пропорционально накоплению в крови (мочевина, сульфаты и др.), называются непороговыми, так как для них порога выведения не существует.
Переносчики апикальной мембраны для симпорта натрия и аминокислот лишь относительно специфичны, каждый из них способен транспортировать несколько видов аминокислот. Так, для глутамата и аспартата существует один вид переносчика, для аргинина, лизина — другой.
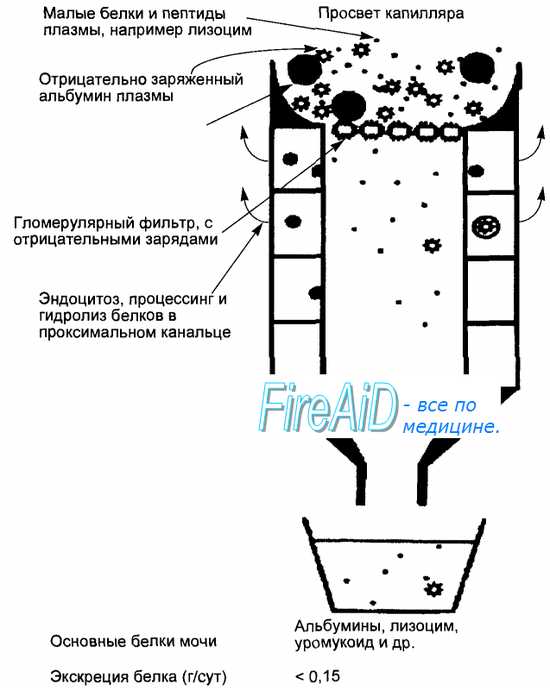
Рис. 14.10. Фильтрация белка и его реабсорбция в проксимальном отделе канальцев.
Гломерулярный фильтр пропускает в первичную мочу лишь мелкие молекулы белков и пептидов. В проксимальных канальцах эти молекулы поглощаются клетками эпителия и гидролизу-ются. Аминокислоты и мелкие пептиды реабсорбируются далее в кровь. С конечной мочой из организма выделяется не более 0,15 г белка в сутки.
Реабсорбция пептидов и белков осуществляется практически полностью в проксимальных канальцах. Количество профильтровавшегося белка относительно невелико и составляет около 1,8 г в сутки. Некоторую его часть составляют альбумины, но фильтрационный барьер клубочков проходят и полипептиды меньшего размера, и в первичную мочу фильтруются, например, соматотропин, а также лизоцим и др. В конечную мочу поступает не более 0,15 г белка в сутки (рис. 14.10). Молекулы альбуминов после связывания с рецепторами на люминальной мембране клеток эпителия канальцев подвергаются эндоцитозу, внутриклеточные пузырьки поглощенного белка сливаются с лизосомами и белковые молекулы гидролизуются пеп-тидазами (аналог внутриклеточного пищеварения). Продукты гидролиза, в основном аминокислоты, выводятся в интерстициальную жидкость и поступают в перитубулярные капилляры. Пептиды, особенно с короткой цепью, подвергаются гидролизу ферментами щеточной каемки (аналог мембранного пищеварения), образующиеся аминокислоты реабсорбируются из просвета канальцев.