Механизм ассиметрии клетки через динамику микротрубочек и моторов
• Асимметрия клетки создается совместным функционированием динамики микротрубочек и моторов
• При обеспечении двигательной активности клетки и позиционировании веретена деления микротрубочки работают вместе с актиновым цитоскелетом
Положение внутриклеточных органелл и общая форма клетки в целом часто характеризуются четко выраженной асимметрией. Способность клетки к созданию двух различных участков, отличающихся друг от друга по расположению и ориентировке органелл и специфических участков цитоплазмы, является фундаментальным и крайне важным свойством. Хотя изолированные клетки могут существовать без этой способности, такие клетки не обладают подвижностью и не могут образовывать специализированные клетки специфической формы, необходимые для создания и существования многоклеточного организма. Например, фибробласты и другие подвижные клетки с одного конца должны расширяться, а с другого — сокращаться, с тем чтобы передвигаться в организме и реагировать на повреждения и инфекции. Концы каждой клетки должны обладать различной активностью и функционировать скоординированным образом.
Асимметричная организация клетки обычно зависит от расположения микротрубочек, их динамики и движения по ним белковых моторов. В организации внутренней архитектуры клетки также участвуют актин и промежуточные филаменты, и все три системы филаментов связаны между собой и контролируют свойства друг друга. В настоящем разделе мы рассмотрим несколько примеров функционирования компонентов цитоскелета микротрубочек, в результате чего возникает клеточная асимметрия. Также будут рассмотрены случаи, иллюстрирующие совместное функционирование микротрубочек и актинового цитоскелета.
В процессе развития, при формировании головного мозга, каждая нервная клетка образует выросты, называемые аксонами, которые контактируют с нейронами-мишенями, образуя синапсы; при этом между нейронами возникает взаимодействие. На конце каждого аксона находится конус роста. Это чрезвычайно подвижная структура, богатая актином и микротрубочками, которая ползет по субстрату вперед. За счет конуса роста происходит удлинение аксона. Во время движения, в соответствии с получаемыми сигналами, конус роста периодически меняет направление. Проделав серию поворотов, он обнаруживает мишень. После этого движение прекращается и образуется синапс.
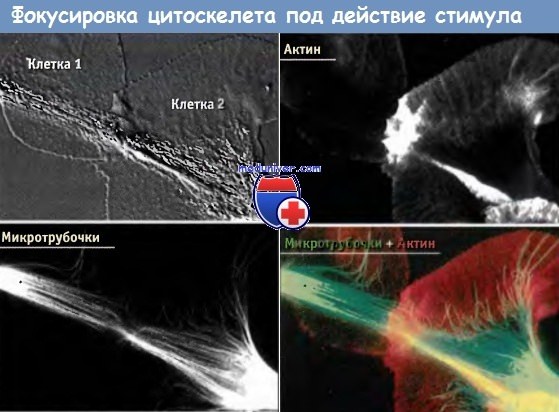
Микротрубочки цитоскелета играют критическую роль в механизме поворотов при перемещении конуса роста. Последний представляет собой крупную уплощенную структуру, способную распластываться по поверхности, по которой происходит движение. В отсутствие внешних сигналов направления движения происходит нуклеация динамичных микротрубочек, расположенных позади конуса роста, причем они растут и сжимаются веерообразным образом. Когда конус роста встречает сигнал, ответная реакция генерируется только на небольшом участке его плазматической мембраны, который контактирует с сигналом. Как показано на рисунке ниже, при этом происходит интересный процесс.
Микротрубочки в пределах этого участка начинают расти по направлению к источнику сигнала. Хотя МАР, обусловливающие этот рост, неизвестны, возможно, что локальная активация МАР сигналом способствует пребыванию микротрубочек в фазе роста в течение длительного времени и приводит к их накоплению в сайте, где возникает сигнал. На рисунке ниже представлены кадры видеоизображения микротрубочек в двух конусах роста через несколько минут после того, как между ними возник контакт. После переориентации и элонгации микротрубочек плюс-направленные кинезины транспортируют по ним везикулы на клеточную периферию. Это приводит к слиянию везикулы с плазматической мембраной и вызывает небольшое локальное расширение клетки. В результате в структуре конуса роста возникает локальная асимметрия. Затем, под действием актинового цитоскелета, мембрана смещается вперед, в направлении хода микротрубочек.
Кадры видеосъемки, которые показывают конусы роста в культуре нейронов.
Процессы, происходящие в конусах роста, когда они входят во взаимный контакт, очень близки к таковым in vivo,
когда конус роста обнаруживает сигнал на другой клетке.
Каждый конус быстро направляет микротрубочки в точки контакта.
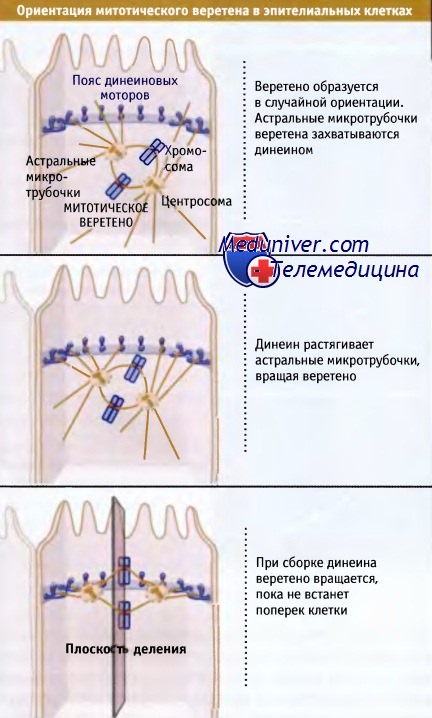
Еще один процесс создания асимметричной организации клетки, в котором участвуют микротрубочки и моторы, представляет собой позиционирование митотического веретена в эпителиальных клетках. При образовании новых клеток, необходимых для роста и восстановления эпителия, необходимо, чтобы делящиеся клетки слоя эпителия образовывали две дочерние клетки одинаковой удлиненной формы и ориентированные таким же образом, как уже существующие. Клетка всегда делится перпендикулярно по отношению к веретену, и поэтому для того, чтобы деление происходило вдоль длинной оси эпителиальной клетки, до разделения хромосом веретено должно быть ориентировано соответствующим образом. Однако после своего образования веретено находится в случайной ориентации. Для того, чтобы приобрести правильную ориентацию, веретено вращается с помощью астральных микротрубочек (микротрубочки на полюсах веретена). Эти микротрубочки высокодинамичны и постоянно ищут периферию клетки. Цитоплазматический динеин прикреплен к местам на плазматической мембране наподобие пояса, который проходит по середине клетки.
Астральные микротрубочки, которые на своем пути встречают этот пояс, связываются с комплексом динеин/динактин, который генерирует тянущее усилие, в результате чего направление веретена приобретает правильную ориентацию.
Наиболее наглядным примером, который иллюстрирует, каким образом возникает клеточная асимметрия, служит деление дрожжей Saccharomyces cerevisiae. Эти дрожжи используются для выпечки хлеба и варки пива.
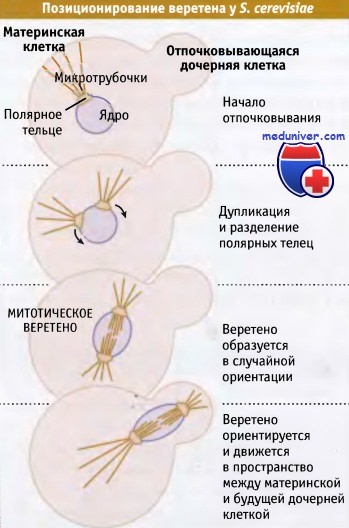
Рисунок иллюстрирует, что дрожжи делятся за счет процесса почкования, при котором небольшой участок поверхности клетки («материнской» клетки) образует вырост, напоминающий почку. В ходе клеточного цикла вырост расширяется и в конце концов становится дочерней клеткой. Примерно в то же время, когда начинается процесс почкования, дуплицируется единственный центр организации микротрубочек в материнской клетке — структура, называемая полярным тельцем, которая входит в ядерную мембрану. Образуются два полярных тельца, которые разделяются, и движутся к противоположным концам ядра, как показано на рисунке ниже.
При делении дрожжей их ядерная оболочка не разрушается, и веретено образуется внутри ядра. В образовании веретена участвуют микротрубочки, нуклеированные двумя полярными тельцами, которые также участвуют в образовании микротрубочек с внешней стороны ядра. Образованное веретено располагается случайным образом, и локализация почки не влияет на его ориентацию.
Все эти процессы происходят в материнской клетке. Для того чтобы обеспечить наследование хромосомного набора обеими клетками, веретено должно быть расположено вдоль оси между материнской клеткой и почкой и ориентировано так, чтобы находиться в пространстве между ними.
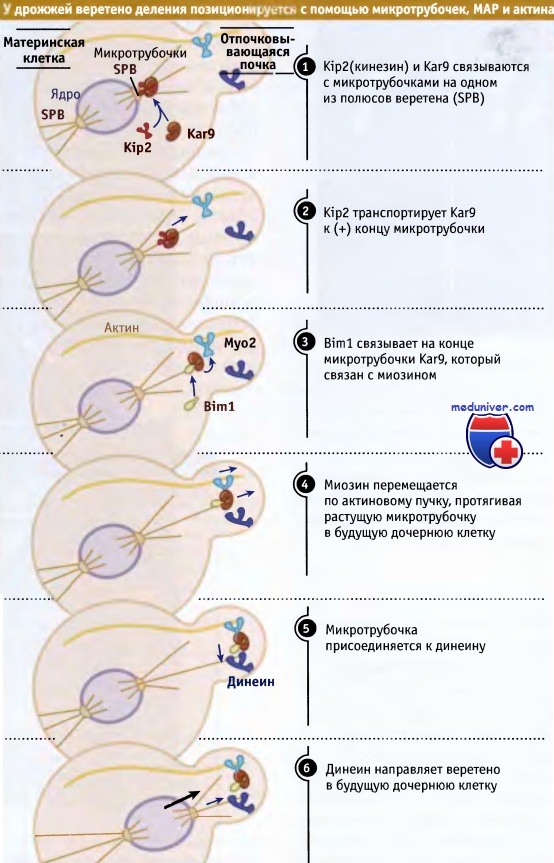
Ориентирование и передвижение веретена происходят с участием микротрубочек, которые из одного полярного тельца выходят в цитоплазму. Каким образом это происходит, показано на рисунке ниже. На одном из двух полярных телец с микротрубочками связывается белок Kar9. От полярного тельца он транспортируется по микротрубочкам в направлении плюс-концов представителем семейства кинезинов (Kip2). Когда Kip2 достигает конца микротрубочки, Kar9 остается там, связываясь с белком +TIP (Bim1). В некоторые моменты времени Kar9 также связывается с высокопроцессивной формой миозина (который относится к семейству миозинов V, способных транспортироваться на большие дистанции вдоль актиновых филаментов). При этом формируется связь между кончиком цитоплазматической микротрубочки и актиновым филаментом. В клетках дрожжей актиновые филаменты представляют собой многонитчатые тяжи, которые через цитоплазму материнской клетки направляются в отпочковывающуюся область, где все тяжи сходятся в одной точке и соединяются с кортексом этой области.
Все актиновые филаменты, составляющие такой пучек, имеют одинаковую полярность, так что он представляет собой полярную структуру, которая указывает направление от материнской клетки к будущей дочерней. Миозин продвигает концы микротрубочек вдоль актинового кабеля, приводя их в отпочковывающуюся область. Попав туда, микротрубочки присоединяются к моторам, закрепленным в слое кортекса, которые движутся к минус-концу (динеиновые и некоторые кинези-новые моторы). Моторы тянут микротрубочки и контролируют состояние их концов при деполимеризации. Таким образом, комбинируя активное протягивание и контролируемую разборку микротрубочек, веретено транспортируется по цитоплазме материнской клетки в будущую дочернюю клетку.
Динамика микротрубочек играет основную роль в ориентировании и позиционировании веретена как в клетках дрожжей, так и эпителия. В обоих случаях существует одна общая черта, состоящая в том, что концы микротрубочек должны найти свое место в слое клеточного кортекса. Однако пути использования для этой цели динамических свойств микротрубочек различаются коренным образом. У дрожжей микротрубочки гораздо более стабильны и не находят мишени по механизму поиска и захвата, который предполагает, чтобы микротрубочки постоянно росли и разбирались. У дрожжей, скорее, цитоплазматические микротрубочки приводят к мишеням.
Динамические свойства начинает проявляться, когда конец микротрубочки транспортируется молекулой миозина вдоль актинового филамента. По мере продвижения конца микротрубочки к нему должны добавляться субъединицы, однако сам он не должен связываться с разорванными актиновыми филаментами. Это становится возможным благодаря белку Bim1. По-видимому, в клетках эукариот этот белок обеспечивает карго возможность находиться на конце растущей микротрубочки.
Если бы цитоплазматические микротрубочки от обоих полярных телец связывались с актином, то позиционирования веретена в клетках дрожжей, очевидно, не происходило бы. В этом случае веретено оставалось бы в материнской клетке, и ось его располагалась перпендикулярно оси между материнской клеткой и почкой. Такая ситуация предотвращается механизмом дупликации полярных телец. Подобно центриолям, старые и новые полярные тельца различаются после каждого раунда дупликации. Так же как и для центросом, образующихся вокруг материнской и дочерней центриолей, состав старых и новых полярных телец различается. В дрожжах это используется для того, чтобы обеспечить присутствие белков, которые избирательно инактивируют Kar9 на новом полюсе веретена. В результате этот белок может связываться только с теми микротрубочками, которые выходят из первоначального, старого полюса. Таким образом, только микротрубочки из старого полюса проходят в будущую дочернюю клетку.
Актин и микротрубочки в двух конусах роста через несколько минут контакта.
Отметьте, что микротрубочки сфокусированы в точке контакта; также там наблюдается массивная центрированная полимеризация актина.
Представлена одна из многих клеток, находящихся в слое эпителия, клетки которого расположены слева направо.
Расположение динеина в виде кольца вокруг клетки обеспечивает конечную ориентацию веретена, независимо от того, как он был ориентирован при образовании.
Эта ориентация обеспечивает деление клетки сверху вниз, создавая две новых клетки в пласте.
Динеин локализуется в месте плотного контакта, где примыкающие клетки в слое контактируют друг с другом.
На трех верхних рисунках показаны клетки, находящиеся на последовательных стадиях клеточного цикла (слева направо).
По мере прохождения цикла отпочковывающиеся участки увеличиваются в размере до тех пор, пока не достигнут размеров материнской клетки.
На нижней фотографии представлена клеточная ДНК. На фотографиях в середине и внизу справа показано, что ДНК можно обнаружить,
только когда дочерняя клетка достигнет размера материнской.
При делении клеток почкующихся дрожжей целостность ядер не нарушается.
Полярное тело веретена, структура, находящаяся в ядерной оболочке,
вызывает нуклеацию микротрубочек в цитоплазме и внутри ядра, где они образуют митотическое веретено.
Веретено образуется в материнской клетке и должно позиционироваться в пространстве между материнской и будущей дочерней клетками.
Правильное позиционирование веретена требует,
чтобы оно сдвигалось по направлению к образующейся дочерней клетке и было ориентировано вдоль оси между дочерней и материнской клеткой.
Перемещение митотического веретена в отпочковывающуюся клетку является следствием совместного функционирования актина и микротрубочек цитоскелета.
Поляризованные пучки актиновых филаментов направляются из материнской в будущую дочернюю клетку и служат для направления микротрубочек от одного из двух полюсов веретена в нужном направлении.
Оказавшись в будущей дочерней клетке, микротрубочки захватываются динеином.
Поскольку динеин присоединен к мембране, он тянет веретено в будущую дочернюю клетку.
Этот механизм не мог бы реализоваться, если бы белки Kar9, Biml связывались с микротрубочками обоих полярных телец.