Роль факторов роста в патогенезе аденомы (доброкачественной гиперплазии) простаты
Андреев А.А., г. Киев, Украина
Аденома (доброкачественная гиперплазия) простаты является заболеванием, которое наблюдается у большинства мужчин старше среднего возраста. Встречаемость аденомы предстательной железы низкая на четвертом десятилетии жизни, однако распространенность заболевания у мужчин к 90-летнему возрасту составляет практически 100%. Хотя у всех мужчин этого возраста имеются гистологические признаки аденомы предстательной железы, только у половины из них наблюдается макроскопическое увеличение простаты и 25% из них нуждаются в хирургическом лечении для ликвидации обструкции.
Установлено, что возраст и функционирующие яички являются факторами, необходимыми для развития доброкачественной гиперплазии простаты (ДГП). Молекулярные основы развития ДГП окончательно не выяснены. В последние годы большое значение придают факторам роста (ФР) в патогенезе ДГП.
Развитие, рост и поддержание функционирования простаты являются андроген-зависимыми. Кастрация приводит к инволюции простаты, а последующее применение экзогенных андрогенов обеспечивает восстановление первоначальных размеров простаты. Тот факт, что у мужчин, которые неспособны образовывать андрогены вследствие генетических причин или кастрации до полового созревания, не развивается ДГП, указывает, что андрогены играют решающую роль в развитии этого заболевания.
В простате тестостерон превращается в 5-альфа-дигидротестостерон (ДГТ) под влиянием фермента 5-альфа-редуктаза, который локализуется на ядерной мембране. ДГТ является активным внутриклеточным андрогеном и осуществляет свою биологическую активность посредством связывания с ядерными протеиновыми рецепторами андрогенов (РА). Комплекс ДГТ-РА может связываться со специфическими участками ДНК, приводя к стимулированию или ингибированию генов, ответственных за регуляцию процессов клеточного роста или образования секреторных продуктов как простатический специфический антиген (ПСА). Однако эксперименты in vitro на культуре ткани показали, что андрогены не оказывают непосредственного стимулирующего влияния на рост эпителиальных клеток, в то время как определенные ФР прямо стимулируют пролиферацию простатических эпителиальных клеток. Таким образом, создается впечатление, что ДГТ не является непосредственно ответственным за эпителиальный рост. Несмотря на это роль андрогенов значительна в патогенезе ДГП, поскольку ДГТ необходим для роста простаты in vivo и поскольку ДГП не развивается у мужчин, кастрированных до полового созревания.
Является признанным тот факт, что тестостерон, путем его превращения в ДГТ, модулирует процессы простатического клеточного роста через баланс между клеточной пролиферацией и программированной смертью клеток (апоптозом). ДГТ сдерживает апоптоз и поддерживает необходимое количество клеток. Снижение ДГТ и снижение интрацеллюлярных РА приводит к инволюции предстательной железы.
Таким образом, андрогены необходимы, но не являются первично ответственными за инициацию простатической клеточной пролиферации. ДГТ "включает" клеточную пролиферацию посредством индуцирования синтеза митогенных пептидов - ФР.
Пептидные ФР могут быть прямыми медиаторами андрогенного воздействия и могут быть ответственны за посредничество в стромально-эпителиальном взаимодействии. Эти ФР являются частью большой семьи пролиферативных и дифференцирующих регуляторных факторов и, как было показано, находятся под регуляторным воздействием андрогенов.
По следующим причинам пептидные ФР признаны локальными факторами, ответственными за опосредование мезенхимальных и эпителиальных взаимодействий, что определяет рост и развитие простаты:
1. Пептидные ФР экспрессированы как в мезенхимальном, так и в эпителиальном компоненте развивающейся простаты.
2. Изолированные простатические эпителиальные и стромальные клетки отвечают на пептидные ФР in vitro.
3. Простатические мезенхимальные и эпителиальные клетки экспрессируют специфические мембранные рецепторы для пептидных ФР.
4. Нейтрализующие антитела для пептидных ФР или их рецепторов ингибируют биологический эффект ФР.
5. Андрогены прямо влияют на экспрессию пептидных ФР простатических клеток.
6. Продукция андрогенов модулируется пептидными ФР.
ФР могут стимулировать или сдерживать клеточную пролиферацию, они активно вовлечены в контроль механизма нормального клеточного роста. Очевидно, что гомеостаз требует равновесия между биологическими эффектами стимулирующих и ингибирующих рост факторами и дисбаланс ФР может явиться причиной ненормального избыточного роста, что наблюдается, в частности, при ДГП. Более того, если гены, кодирующие эти протеины, гиперактивны или мутированы так, что процесс их регулирования нарушен, может последовать неконтролируемый рост, характерный для рака.
Классически, эндокринный эффект - это когда гормоны синтезируются железой внутренней секреции и транспортируются к клеткам-мишеням через циркуляцию. ФР действуют посредством паракринных, аутокринных и, вероятно, интракринных эффектов. При паракринном эффекте ФР продуцируются одной клеткой и диффундируют к другой клетке, обеспечивая его биологическое действие, тогда как при аутокринном эффекте клетка использует ФР, который сама продуцирует и выделяет. При интракринном эффекте фактор остается внутри клетки.
Недавние исследования наводят на мысль, что интрапростатический ДГТ образуется в стромальных и базальных эпителиальных клетках. Таким образом, эффект ДГТ на эпителиальные клетки скорее паракринный, чем аутокринный. Более того, предварительные доказательства наводят на мысль, что ДГТ, образующийся в периферических тканях, таких как кожа и печень, - может оказывать истинный эндокринный эффект на простату.
В настоящее время очевидно, насколько важно биологическое взаимодействие между простатической стромой и эпителием. В ряде исследований было показано, что во время эмбрионального развития простаты наблюдаются индуктивные сигнальные процессы между стромой и эпителием. Под влиянием андрогенов и в некоторой степени эстрогенов стромальные клетки продуцируют факторы, которые регулируют рост и дифференцировку эпителиальных клеток. И наоборот, эпителиальная ткань оказывает влияние на поведение стромального компонента. Исследования поддерживают эту концепцию тесного взаимодействия между стромой и эпителиальными элементами с индуктивными сигналами между составляющими, контролируемыми ФР, включая рост, дифференцировку и, возможно, функционирование. Дисбаланс этих сложных взаимодействий между двумя составляющими может иметь отношение к патогенезу ДГП.
Начальный рост и дифференцировка простаты и рост железы после полового созревания непрямым образом усиливаются андрогенами через андроген-зависимые ФР стромального происхождения. Предполагается, что при достижении железой взрослого размера, факторы, сдерживающие рост, предотвратят дальнейший рост простаты. Гомеостаз поддерживается посредством баланса между рост-стимулирующими и ингибирующими факторами.
Семейство эпидермальных факторов роста (ЭФР) является универсальным стимулятором для эпителиально-клеточного роста. Высокие концентрации ЭФР определяли в секрете простаты, пролиферация простатических эпителиальных клеток может стимулироваться ЭФР. Экспрессия рецепторов ЭФР в эпителиальных клетках была продемонстрирована иммунологически и подтверждена в основном для базальных клеток. Не совсем ясно, отличаются ли уровни ЭФР и рецепторов ЭФР в нормальной, гиперплазированной и опухолевой ткани. Исследователи не наблюдали различий между уровнями ЭФР в экстрактах простатической ткани, полученной у пациентов с раком и доброкачественной гиперплазией простаты.
Семейство ЭФР представлено 5 членами, активирующими рецепторы ЭФР: ЭФР, трансформирующий фактор роста альфа (ТФР-альфа), амфирегулин, гепарин-связывающий ЭФР-подобный ФР (ГС-ЭФР) и бетацеллюлин. Вышеперечисленные ФР потенциально вовлечены в регуляцию роста простаты. ЭФР-подобная активность определяется в интактной простате и секрете простаты. ТФР-альфа был определен в человеческой простате иммунологическими и молекулярными методами. Амфирегулин определен как продукт человеческих простатических эпителиальных клеток и его экспрессия в культуре регулируется андрогенами.
ГС-ЭФР был идентифицирован как продукт гладкомышечных клеток простаты in vivo. ГС-ЭФР связывает и активирует рецепторы ЭФР подобно ЭФР и ТФР-альфа; однако, было показано, что он является таким же мощным митогеном для гладко-мышечных клеток как классический митоген гладко-мышечных клеток - тромбоцитарного происхождения фактор роста (ТПФР). Проведенные исследования позволяют предположить, что ГС-ЭФР, синтезированный гладкомышечными клетками простаты, может играть роль в распространении стромального компонента.
Семейство факторов роста фибробластов (ФРФ) представлено тремя членами: кислый ФРФ (кФРФ/ФРФ1), основной ФРФ (оФРФ/ФРФ2), фактор роста кератиноцитов (ФРК/ФРФ7). оФРФ/ФРФ2 является плеотропным полипептидом, способным стимулировать пролиферацию различных культур клеток, включая фибробласты простаты человека. оФРФ/ФРФ2 также вызывает рост эндотелиальных клеток in vitro, инициирует неоваскуляризацию in vivo и является важным опухолевым фактором ангиогенеза. оФРФ/ФРФ2 определяется в значительном количестве в нормальной человеческой простате и секрете простаты.
Существуют доказательства того, что ФРФ принимают участие в развитии ДГП. Применяли радиоиммунный метод для измерения уровней оФРФ/ФРФ2 в нормальной и гиперплазированной простатах. Гиперплазированные железы характеризовались 2-3 кратным увеличением этого ФР в сравнении с гистологически неизмененными простатами.
ФРК/ФРФ7 был идентифицирован как продукт простатических стромальных клеток. Этот фактор, продуцируемый простатическими фибробластами в культуре клеток андрогензависимым образом, является митогенным в культуре нормальных простатических эпителиальных клеток. Рост-стимулирующмй эффект ФРК/ФРФ7 в культуре эпителиальных клеток может быть отменен добавлением трансформирующего фактора роста бета (ТФР-бета), что указывает на то, что оба ФР вовлечены в регуляцию эпителиальной пролиферации.
ТФР-бета является самым важным физиологическим ингибитором пролиферации эпителиальных клеток. ТФР-бета-1 также регулирует биосинтез и деградацию экстрацеллюлярного матрикса, является мощным иммуносупрессором и стимулирует клетки к апоптозу. Раковые клетки, включая простатические опухолевые клетки, часто теряют способность отвечать на ТФР-бета-1. Трофический эффект тестостерона и ДГТ может ингибироваться добавлением ТФР-бета-1 к культуральной среде. Блокирующий эффект носит дозо-зависимый характер. ТФР-бета играет роль в ограничении дуктальной пролиферации и арборизации также как в стимулировании гладко-мышечного роста. Гиперэкспрессия ТФР-бета-1 нарушает развитие простаты путем снижения дуктального ветвления и увеличения гладких мышц вокруг ацинарных протоков. В основном ТФР-бета считается ингибитором эпителиально-клеточного роста. В простате ТФР-бета также действует как ингибитор пролиферативных процессов и ответственен за апоптоз. Таким образом, ТФР-бета может регулировать простатический эпителиально-клеточный рост путем сдерживания рост-стимулирующих эффектов ЭФР и ФРФ. Утрата чувствительности эпителиальных клеток к ТФР-бета или другим свойствам ТФР-бета, включая стромально-клеточную пролиферацию, может способствовать развитию ДГП.
Другие ФР, которые вероятно важны в гомеостазе простаты - члены семейства инсулиноподобных факторов роста (ИФР). Культура простатических эпителиальных клеток стимулируется очищенными ИФР1 и ИФР2. ИФР1 и ИФР2 не определяются в среде простатических эпителиальных клеток, что указывает на то, что аутокринная регуляция маловероятна. Недавние исследования показали, что культура простатических фибробластов продуцирует ИФР2, что может свидетельствовать о существовании паракринной регуляции простатического эпителиального роста. Другие протеины, которые вовлечены в митогенный эффект ИФР - ИФР-связывающие белки. Точный механизм действия их остается неясным, но вероятно они вовлечены в контроль доступности ИФР для взаимодействия их с рецепторами.
Мощными митогенами для фибробластов и гладкомышечных клеток простаты являются изоформы ТПФР. ТПФР обеспечивает пролиферацию большинства клеток фибромускулярной простатической стромы (недифференцированных фибробластов и гладкомышечных клеток). Было показано, что стромальные клетки, выделенные из человеческой простатической ткани, экспрессируют высокоаффинные рецепторы для изоформ ТПФР и демонстрируют увеличение пролиферативной активности в ответ на экзогенный ТПФР.
Фактор роста нервов (ФРН)-подобный протеин был идентифицирован в среде человеческих простатических стромальных и неопластических эпителиальных клеток и был локализован иммуноцитохимически в стромальном компоненте нормальных и гиперплазированных простат и простатической аденокарциноме. ФРН-рецепторы локализуются преимущественно в эпителии, что наводит на мысль, что ФРН-подобные полипептиды могут быть стромально-обусловленными медиаторами эпителиально-клеточной пролиферации в человеческой простате.
Потенциально важный компонент простатической стромы, не изученный детально, представлен воспалительно-клеточными инфильтратами, включающими лимфоциты и макрофаги моноцитарного происхождения. Имеются сообщения об интенсивной инфильтрации ткани гиперплазированной простаты хронически активированными Т-клетками. Отсутствие гранулоцитов в этих инфильтратах наводит на мысль, что аккамуляция мононуклеарных клеток в простате маловероятно является результатом инфекции. Т-клетки периферической крови способны образовывать и секретировать три гепарин-связывающих ФР: ГС-ЭФР, оФРФ/ФРФ2, а также сосудистый эндотелиальный фактор роста (СЭФР) - мощный ангиогенный фактор и эндотелиальный митоген. оФРФ/ФРФ2 также является эндотелиальным митогеном и ангиогенным фактором. СЭФР является продуктом Т-клеток, инфильтрирующих человеческую простату. Т-клетки обладают способностью секретировать мощные эпителиальные, сосудистые и стромальные митогены в интерстициальную ткань, таким образом, что этот процесс не зависит от иммунорегуляторных сигналов. В контексте ДГП это наводит на мысль, что Т-клеточные инфильтраты не функционируют как часть иммунной системы, но представленные в значительном количестве, могут способствовать стромальной и железистой гиперплазии.
Исследователи для изучения влияния ФР на рост простаты использовали простатические стромальные (ПС) и простатические эпителиальные (ПЭ) клеточные культуры из нормальных и гиперплазированных человеческих простат. Стромальные клетки синтезировали оФРФ/ФРФ2 и КФР/ФРФ7. оФРФ/ФРФ2 являлся мощным митогеном для ПС, но слабым митогеном для ПЭ, в то время как КФР/ФРФ7 являлся митогеном только для ПЭ. И ПС и ПЭ синтезировали ТФР-бета. ТФР_бета ингибировал пролиферацию как ПС так и ПЭ. Эти наблюдения наводят на мысль, что рост простатической стромы регулируется взаимодействием ЭФР, ФРФ и ТФР-бета. Рост простатического эпителия позитивно регулируется ЭФР и КФР и негативно ТФР-бета. В нормальной простате уровень клеточной пролиферации соответствует уровню клеточной смерти. Это подразумевает, что влияние оФРФ/ФРФ2 соответствует влиянию ТФР-бета в строме, в то время как влияние ЭФР и КФР/ФРФ7 соответствует влиянию ТФР-бета в эпителии. В ДГП, характеризующейся стромальной гиперплазией, влияние оФРФ/ФРФ2 выше ТФР-бета, в то время как в эпителиальной гиперплазии влияние КФР/ФРФ7 и ЭФР превосходит таковое ТФР-бета.
Возрастание понимания того факта, что пептидные ФР играют важную роль в патогенезе ДГП, вместе с расшифровкой путей, по которым взаимодействие клеток стромы и эпителия воздействуют на продукцию ФР, обеспечивают новое фармакологическое объяснение и обоснование лечения ДГП ингибиторами ФР. Этот подход предполагает возможность предохранения и сохранения единства клеточного состава предстательной железы и, таким образом, нормализации как ее структуры, так и функции.
Андрей Александрович Андреев
Зав. отд. урологии Центрального госпиталя МВД Украины

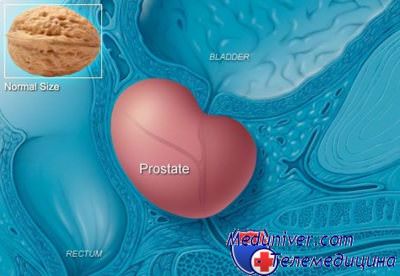
 Аденома (доброкачественная гиперплазия) простаты является заболеванием, которое наблюдается у большинства мужчин старше среднего возраста. Встречаемость аденомы предстательной железы низкая на четвертом десятилетии жизни, однако распространенность заболевания у мужчин к 90-летнему возрасту составляет практически 100%. Хотя у всех мужчин этого возраста имеются гистологические признаки аденомы предстательной железы, только у половины из них наблюдается макроскопическое увеличение простаты и 25% из них нуждаются в хирургическом лечении для ликвидации обструкции.
Аденома (доброкачественная гиперплазия) простаты является заболеванием, которое наблюдается у большинства мужчин старше среднего возраста. Встречаемость аденомы предстательной железы низкая на четвертом десятилетии жизни, однако распространенность заболевания у мужчин к 90-летнему возрасту составляет практически 100%. Хотя у всех мужчин этого возраста имеются гистологические признаки аденомы предстательной железы, только у половины из них наблюдается макроскопическое увеличение простаты и 25% из них нуждаются в хирургическом лечении для ликвидации обструкции.
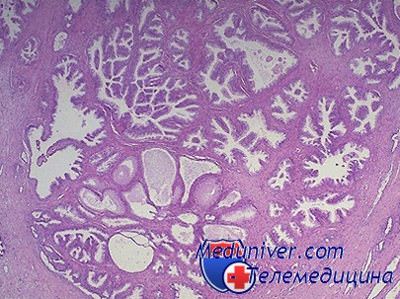
 ФР могут стимулировать или сдерживать клеточную пролиферацию, они активно вовлечены в контроль механизма нормального клеточного роста. Очевидно, что гомеостаз требует равновесия между биологическими эффектами стимулирующих и ингибирующих рост факторами и дисбаланс ФР может явиться причиной ненормального избыточного роста, что наблюдается, в частности, при ДГП. Более того, если гены, кодирующие эти протеины, гиперактивны или мутированы так, что процесс их регулирования нарушен, может последовать неконтролируемый рост, характерный для рака.
ФР могут стимулировать или сдерживать клеточную пролиферацию, они активно вовлечены в контроль механизма нормального клеточного роста. Очевидно, что гомеостаз требует равновесия между биологическими эффектами стимулирующих и ингибирующих рост факторами и дисбаланс ФР может явиться причиной ненормального избыточного роста, что наблюдается, в частности, при ДГП. Более того, если гены, кодирующие эти протеины, гиперактивны или мутированы так, что процесс их регулирования нарушен, может последовать неконтролируемый рост, характерный для рака.
 ГС-ЭФР был идентифицирован как продукт гладкомышечных клеток простаты in vivo. ГС-ЭФР связывает и активирует рецепторы ЭФР подобно ЭФР и ТФР-альфа; однако, было показано, что он является таким же мощным митогеном для гладко-мышечных клеток как классический митоген гладко-мышечных клеток - тромбоцитарного происхождения фактор роста (ТПФР). Проведенные исследования позволяют предположить, что ГС-ЭФР, синтезированный гладкомышечными клетками простаты, может играть роль в распространении стромального компонента.
ГС-ЭФР был идентифицирован как продукт гладкомышечных клеток простаты in vivo. ГС-ЭФР связывает и активирует рецепторы ЭФР подобно ЭФР и ТФР-альфа; однако, было показано, что он является таким же мощным митогеном для гладко-мышечных клеток как классический митоген гладко-мышечных клеток - тромбоцитарного происхождения фактор роста (ТПФР). Проведенные исследования позволяют предположить, что ГС-ЭФР, синтезированный гладкомышечными клетками простаты, может играть роль в распространении стромального компонента.
 Фактор роста нервов (ФРН)-подобный протеин был идентифицирован в среде человеческих простатических стромальных и неопластических эпителиальных клеток и был локализован иммуноцитохимически в стромальном компоненте нормальных и гиперплазированных простат и простатической аденокарциноме. ФРН-рецепторы локализуются преимущественно в эпителии, что наводит на мысль, что ФРН-подобные полипептиды могут быть стромально-обусловленными медиаторами эпителиально-клеточной пролиферации в человеческой простате.
Фактор роста нервов (ФРН)-подобный протеин был идентифицирован в среде человеческих простатических стромальных и неопластических эпителиальных клеток и был локализован иммуноцитохимически в стромальном компоненте нормальных и гиперплазированных простат и простатической аденокарциноме. ФРН-рецепторы локализуются преимущественно в эпителии, что наводит на мысль, что ФРН-подобные полипептиды могут быть стромально-обусловленными медиаторами эпителиально-клеточной пролиферации в человеческой простате.