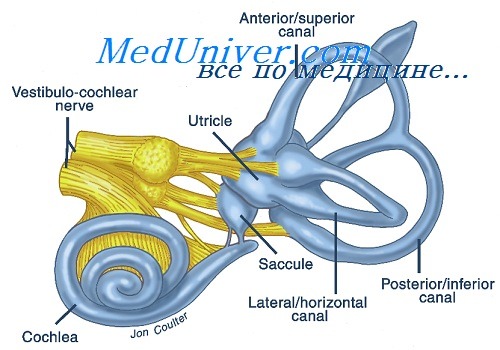
Звуковой анализатор - улитка. Строение звуковоспринимающего аппарата
В улитке совершается дальнейшая доставка звуков к рецептору, а в нем происходит трансформация звуковой энергии в процесс нервного возбуждения. Поэтому в отношении процессов, совершающихся в улитке, следует раздельно различать функции звукопроведения и звуковосприятия.
В звукопроведении принимают участие как жидкие среды, так и мембраны перепончатого лабиринта. Точное изучение этих физических процессов началось лишь недавно, а прежние теории слуха базировались либо на теоретических соображениях, либо на несовершенных экспериментальных моделях, так как благодаря плотной лабиринтной капсуле и малой величине процессы в улитке были недоступны для методов прямого наблюдения.
Благодаря разработке микрооперации, применению стробоскопического наблюдения под микроскопом, а также использованию электрофизиологических методов, колебательные процессы в улитке изучены с большей полнотой.
Классическая теория Гельмгольца, согласно которой звук определенной частоты вызывает соколебания определенного участка базилярной мембраны по законам механического резонанса, в настоящее время дополнена теориями, придающими большее значение гидродинамическим процессам в улитке.
Согласно законам инерции, более частые колебания способны сообщить свой ритм только небольшому столбу жидкости; очень медленные колебания вызывают сдвиги большого столба лимфы. Кроме того, более частые колебания ведут к значительному повышению трения в скалах, ввиду чего изгиб мембран происходит в участках, ближе лежащих к овальному окну, которые благодаря малой ширине и сильной натянутости изгибаются труднее, при медленных же колебаниях изгибается широкая и менее натянутая мембрана у верхушки улитки, так как для этого требуется меньшая сила.
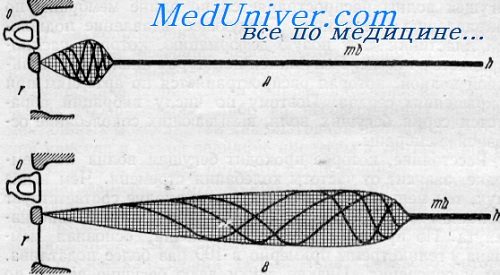
Следующее вдавление подножной пластинки дает новую деформацию, которая распространяется так же. Получается некоторая аналогия с пульсовой волной, которая распространяется по артериям при сокращениях сердца. Поэтому но числу вибраций образуется серия бегущих волн, вызывающих соколебание основной мембраны.
Не трудно видеть, что при низких звуках количество раздражаемых нервных элементов весьма велико. Максимума амплитуды бегущая волна достигает непосредственно перед местом погашения волны, т. е. после достижения максимума амплитуды деформация быстро падает до нуля. При сильных звуках в этом участке могут образоваться вихревые движения лимфы.
Следует предположить, что ощущение высоты звука зависит от месторасположения максимального изгиба основной мембраны.
Таким образом, согласно всем теориям, звуки определенной высоты раздражают определенные участки основной мембраны (place theory), что и объясняет способность уха различать частоты и осуществлять разложение сложных звуков (пространственная теория).
Таким образом, установлена способность улитки к первичному анализу. При помощи условнорефлекторной методики (Л. А. Андреев) подтверждено, что у собаки область восприятия высоких тонов находится у основания, а низких — у верхушки улитки. Такие же результаты были получены при помощи метода регистрации токов улитки. При этом разрушение отдельных участков улитки производилось у кошек, кроликов и морских свинок [Девис, Крейчи (Davis, Krejci), В. Ф. Ундриц и др.].
В наших опытах мы разрушали второй завиток улитки морской свинки и отводили токи с круглого окна, причем обнаружили явное падение потенциалов на средние тона. Однако токи обнаруживались, хотя и в ослабленном виде, и на все другие тона. В свете новой теории бегающих волн это делается понятным, так как клеточные элементы основного завитка раздражаются при любом звуке. Этим, вероятно, отчасти объясняется большая ранимость этой зоны при акустической травме и при возникновении профессиональной тугоухости. О пространственном расположении восприятия в улитке говорят также опыты отведения потенциалов при помощи микроэлектродов с отдельных завитков.
Число нервных элементов вполне допускает различение высоты звука по пространственному принципу. Исходя из величины разностных порогов частоты, наше ухо отличает около 1500 раздельно воспринимаемых частот. Если принять длину основной мембраны за 30 мм, то на каждом миллиметре расположилось бы 50 частот. При этом каждая частота отстояла бы от соседа на расстоянии 0,02 мм. На таком участке расположено около 20 волосковых клеток.
Трудности возникают по другому поводу: по любой из новых пространственных теорий, деформация основной перепонки под влиянием определенного тона занимает гораздо большую зону, чем 0,02 мм. Поэтому для объяснения тонкого различения ухом высоты звука приходится прибегать к добавочным гипотезам, которые учитывают особенности нервного раздражения, причем особая роль в этом отношении принадлежит центральным звеньям анализатора — проводникам и центрам.
Бегущая волна (по М. Portniann).
А — при высоких звуках; В — низких частотах; О — овальное окно; Г — круглое окно. mb — основная перепонка; h — геликотрема
Приходится допускать, что при раздражении группы волосковых клеток возбуждение передается только по тем проводникам, которые получают импульсы с участка максимального изгиба. Электрофизиологические исследования центральных звеньев говорят о большом значении процессов торможения.
Изучение процессов, происходящих при раздражении рецептора (волосковых клеток), электрофизиологическими методами началось 30 лет тому назад, когда в 1930 г. Уивер и Брей (Wever, Bray) сообщили о том, что раздражение улитки звуком сопровождается генерацией электрических потенциалов. Характер тока улитки по силе, частоте и виду в широких пределах соответствовал кривой звука. Эти токи распространяются по законам электропроводимости и легко могут быть обнаружены при контакте отводящего электрода с мембраной круглого окна или слуховым нервом.
Они легко записываются в виде осциллограммы, а также могут быть выслушаны после соответственного усиления при подаче на любой телефон или громкоговоритель. Таким образом, улитка представляет собой живой микрофон, в связи с чем эти потенциалы получили название микрофонных потенциалов улитки. Эти потенциалы представляют собой существенный элемент при раздражении рецептора; при отсутствии их отсутствует и слух. Поэтому они являются замечательным индикатором для изучения нарушений работы рецептора.
Многие данные о функции среднего уха базируются на результатах, полученных путем изучения микрофонных потенциалов (В. Ф. Ундриц и др.); повреждение же центральных звеньев анализатора (например, декортикация животного) не нарушает генерации микрофонных потенциалов, поэтому наличие их еще не доказывает сохранности слуха, а только свидетельствует о функциональной целости улитки.
Микрофонные потенциалы были получены и у человека (при прикладывании электрода к круглому окну через перфорацию барабанной перепонки), но они гораздо слабее, чем у кошки и других экспериментальных животных [Нилен (Nylon), Г. В. Гершуни и др.].
Однако очень скоро было обнаружено (Davis), что потенциалы, отводимые с круглого окна, представляют собой не только микрофонные потенциалы, но к ним примешиваются и другие, а именно акционные потенциалы, получаемые от активной деятельности нервных элементов. Источник акционных потенциалов не вызывает сомнений — он аналогичен акционным токам в других нервах и подчиняется тем же закономерностям. Эти потенциалы очень чувствительны к холоду, недостатку кислорода и т. д.
Они имеют рефрактерный период и одиночное нервное волокно не может дать более 700 импульсов в секунду. Благодаря синхронной деятельности многих волокон (согласно теории залпов) могут наблюдаться и более частые колебания тока. При отведении потенциалов с круглого окна наименьший латентный период имеют микрофонные потенциалы, поэтому для того, чтобы наблюдать акционные потенциалы в чистом виде, применяют очень короткие звуковые сигналы. В этом случае на осциллограмме микрофонные и акционные потенциалы записываются раздельно — раньше идет запись микрофонных потенциалов, а потом акционных.