В понимание нарушений процессов регуляции в опухолевых клетках значительный вклад внесли недавние открытия последовательностей ДНК в генотипах опухолевых и здоровых клеток, которые оказались гомологичными вирусным онкогенам. Термин «онкоген» может быть не самым удачным, так как при упоминании о злокачественной трансформации имеется в виду либо ненормальная активация здоровых генов, либо повреждения генов, ведущие к их гиперэкспрессии.
С этой точки зрения данные гены не являются генами рака, а лишь генами-регуляторами пролиферации. Регуляторная функция последних может быть нарушена, что и ведет к развитию раковой опухоли, включая ее инвазию и метастазирование.
В настоящее время идентифицировано уже множество генов, гомологичных определенным участкам ДНК, кодируемым ретровирусами. Механизмы активации этого класса генов при разных заболеваниях варьируют. Так при заболевании лимфомой Беркита имеет место реципрокная транслокация части хромосомы 8 с хромосомами 2, 14 и 22. Также во многих случаях определяется транслокация хромосом 8 и 14.
Онкоген c-myc расположен в переносимом участке хромосомы 8, и при транслокации происходит реорганизация его структуры. При хронической гранулоцитарной лейкемии (ХГЛ) онкоген с-abl вместе со свом транслокационным партнером, геном bcr, генерирует образование комплексного белка, который вызывает экспрессию фермента тирозиновая киназа — стимулятора деления клеток. В данном случае активация только прото-онкогена в большинстве случаев не ведет к развитию онкологического заболевания.
И конечно, развитие ХГЛ в более агрессивные формы лейкемии требует дальнейших мутаций. Повышенная экспрессия клеточных онкогенов была обнаружена на многих клеточных линиях человеческого рака: это экспрессия гена с-тус в культурах мелкоклеточного рака легкого и лимфомы Беркитта, и гена N-myc при нейробластоме. Очень часто встречаются мутации гена с-KIT при опухолях стромы желудочно-кишечного тракта (ОЖКТ). Этот онкоген гомологичен гену (v-KIT) вируса кошачьей фибросаркомы. Данный ген кодирует производство рецептора к стимулятору клеточного роста, тирозиновои киназе.
Антагонист тирозинкиназы, разработанный для блокировки ее повышенной активности при хронической гранулоцитарной лейкемии, также ингибирует экспрессию гена c-KIT, что ведет к регрессии опухоли.
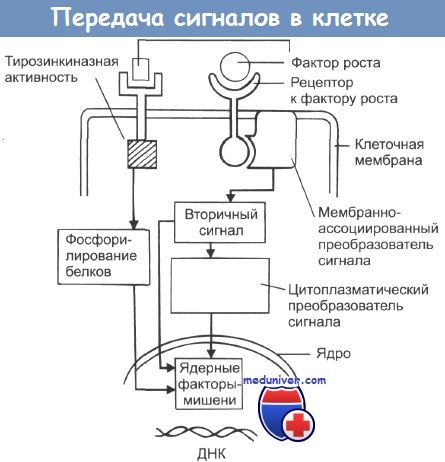
На рисунке показана упрощенная схема событий, происходящих после того, как фактор роста связывается с мембранным рецептором клетки. Рецептор активирует сигнальный белок, который затем может связываться с молекулами гуани-новых нуклеотидов (на этом этапе он получает название G-протеин). Сигнальные молекулы G-протеина стимулируют активность вторичного мессенджера, такого как циклическая аденозин 3', 5'-фосфатаза. При альтернативном механизме активации плазматическая часть рецептора может запускать тирозинкиназную активность, что ведет к фосфорилированию тирозиновых оснований внутриклеточных белков, часть из которых являются стимуляторами клеточного роста и индукторами деления.
Ряд известных к настоящему моменту онкогенов и их производных представлен в таблице. Онкопротеин c-erb В1, являющийся рецептором к ЭФР, принадлежит к классу тирозинкиназных рецепторов (в эту группу также входят HER2/neu (erbB2), erbB3 и erbB4). Связывание ЭФР с рецептором (в качестве лиганда) запускает его тирозинкиназную активность, что ведет к фосфорилированию клеточных белков.
Гиперэкспрессия или мутагенное повреждение комплекса ЭФР-рецептор при раке ведет к усилению рецепторной активности. Основными мишенями для действия комплекса ЭФР-рецептор являются ras-rat-MAP киназы, протеинкиназный каскад реакций Jak/Star, и PI-3-киназный каскад реакций. Действие многих современных лекарственных препаратов направлено как раз на ингибирование таких каскадов реакций, а моноклональные антитела предназначаются для блокировки самого рецептора.
Мембранные белки-рецепторы могут связываться с гуанозин 5'-трифосфатазой (ГТФ), а ГТФ-азная активность приводит к прекращению передачи сигналов от рецепторов. Дефектный G-протеин, кодируемый мутировавшим ras-онкогеном, как раз страдает дефицитом ГТФ-азной активности и не может своевременно прекратить подачу сигналов клетке. Семейство ras-онкогенов состоит из трех членов (H-ras-1, K-ras-2, N-ras) и включает в себя ряд других, частично гомологичных белков {rbo, R-ras, ras, rab). Повышенное число ras-мутаций было выявлено у 40% больных раком толстого кишечника и у 95% больных раком поджелудочной железы. У всех этих больных отмечается повышенная продукция клетками ras-белка.
В культурах клеток in vitro ras-белок вызывает злокачественную трансформацию фибробластов, его экспрессия также ассоциируется с дифференцировкой клеток в процессе эмбрионального развития.