Весь генетический материал клетки находится в упакованном виде в относительно ограниченном объеме. У бактерий генетический материал находится в виде нуклеоида, который образует в клетке дискретную массу. В интерфазном ядре эукариотической клетки генетический материал упакован в массе хроматина.
Упаковка хроматина характеризуется гибкостью и на протяжении клеточного цикла меняется. Во время деления (митоза или мейоза), когда становятся видимыми отдельные хромосомы, интерфазный хроматин становится упакованным более плотно.
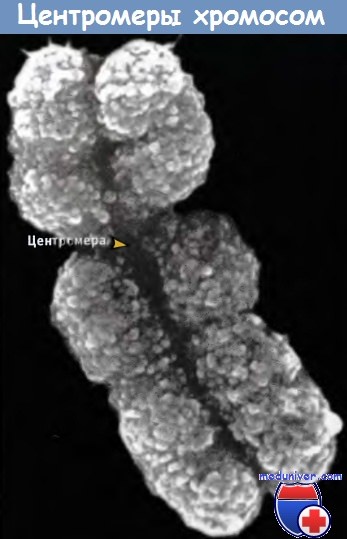
Хромосомы представляют собой форму изоляции генетического материала при клеточном делении. Характерной структурной особенностью, благодаря которой это достигается, является центромера, представляющая собой перетяжку на теле хромосомы, видимую в световом микроскопе. При достаточно высоком разрешении видно, что центромера включает кинетохор, структуру, посредством которой она прикрепляется к микротрубочкам.
Обычно эукариотические хромосомы состоят из очень длинных линейных молекул ДНК, и еще одной их характерной особенностью является наличие теломеры, которая стабилизирует концы хромосом и достраивается по специальному механизму, в обход трудности репликации концов линейной ДНК.
ДНК упакована с высокой плотностью. Для нуклеоида бактерий она составляет около 10 мг/мл, для ядра клетки эукариот порядка 100 мг/мл, а для головки фага Т4 более 500 мг/мл. Растворы такой концентрации представляют собой вязкий гель, и по непонятной причине столь высокая концентрация необходима для проявления белками способности находить сайты связывания на ДНК. Различные процессы, протекающие с участием ДНК, например репликация и транскрипция, должны осуществляться в очень ограниченном пространстве.
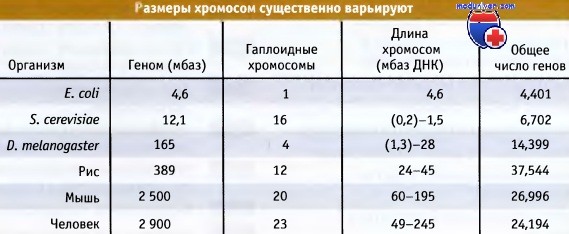
Организация ядерного материала должна соответствовать переходам от неактивного к активному состоянию хроматина. Рисунок ниже иллюстрирует сильные различия в размере генома и в содержании ДНК хромосом некоторых организмов.
Длина линейной молекулы ДНК существенно превышает размер того региона, в котором она находится. Плотно упакованная структура образуется в результате связывания ДНК с основными белками. Положительно заряженные группы белков нейтрализуют отрицательно заряженные группы в ДНК. Организация нуклеопротеидного комплекса определяется взаимодействием белков с ДНК, в результате чего образуется плотно спирализованная структура.
Таким образом, в отличие от распространенной картинки, на которой ДНК представлена в виде протяженной двойной спирали, она, как правило, подвержена структурным деформациям сгиба или скручивания, обеспечивающим образование более компактной структуры.
Большая часть хроматина обладает довольно рыхлой структурой; он называется эухроматин и содержит активные гены. Отдельные участки хроматина упакованы более плотно; они соответствуют генетически неактивному гетерохроматину.
Какова общая структура хроматина и какие существуют различия между активными и неактивными последовательностями? Высокая плотность упаковки генетического материала позволяет предполагать, что ДНК не может быть прямо упакована в конечную структуру хроматина. Должна существовать определенная иерархия ее пространственной организации. Основной вопрос заключается в специфичности упаковки. Следует ли скручивание ДНК определенному общему принципу или же происходит по разному в каждой индивидуальной копии генома? Каким образом меняется вид упаковки при репликации или транскрипции сегмента ДНК?
Основная структурная единица хроматина во всех эукариотических клетках одна и та же. Нуклеосома содержит примерно 200 пн ДНК и является октамером, состоящим из небольших основных белков, имеющим форму бусины. Эти белки представляют собой гистоны. Они образуют внутреннее ядро частицы, а ДНК находится на ее поверхности. В интерфазном ядре и в митотических хромосомах нуклеосомы являются постоянным компонентом эу- и гетерохроматина.
Нуклеосомы обеспечивают начальный уровень организации хроматина. Они обеспечивают упаковку молекулы ДНК длиной 67 нм в структуру, диаметром 11 нм. Компоненты нуклеосом и их строение хорошо изучены. Последовательность нуклеосом, расположенная линейно образует «10-нм нуклеосомную нить».
Следующий уровень организации представляет собой скручивание последовательности нуклеосом в спиральные витки. При этом создается фибрилла диаметром около 30 нм; она присутствует в интерфазном хроматине и в митотических хромосомах. В результате плотность упаковки увеличивается в 6-7 раз на ед. длины. Упаковка в такую структуру требует участия дополнительных белков и исследована недостаточно.
Окончательная плотность достигается на третьем уровне организации, включающем упаковку самой 30-нм фибриллы. Этот уровень приводит к 50-кратно-му увеличению плотности упаковки в эухроматине по сравнению с упаковкой в 30-нм фибрилле. Эухроматин периодически упаковывается в митотические хромосомы, при этом плотность упаковки возрастает в 5-10 раз. Обычно плотность упаковки гетерохроматина такая же, как в митотических хромосомах.
В хроматине содержится примерно в два раза больше белка, чем ДНК. Примерно половина всего количества белка сосредоточена в нуклеосомах. Количество РНК составляет менее 10% от содержания ДНК. Большая часть РНК представлена вновь образованными транскриптами, еще связанными с матричной ДНК.
Изменения структуры хроматина обеспечиваются связыванием дополнительных или модификациями существующих белков. Репликация и транскрипция требуют раскручивания ДНК, и, таким образом, должны включать разупаковку структуры, которая позволяет соответствующим ферментам работать на ДНК. Вероятно, это сопровождается изменениями на всех уровнях организации.
Все белки хроматина, за исключением гистонов, относятся к негистоновым белкам. По сравнению с гистонами, в разных тканях и организмах эти белки характеризуются более разнообразным составом, и в хроматине их содержится меньше, чем гистонов. Они также составляют более многочисленную группу, так что любой индивидуальный негистоновый белок присутствует в количествах гораздо меньших, чем любой гистон.
Пара сестринских хроматид в митозе,
каждая из которых содержит фибриллу (диаметром 30 нм),
компактно упакованную в хромосому.