Взаимодействие белков с гистонами при образовании гетерохроматина
• НР1 представляет собой ключевой белок, образующий гетерохроматин. Он действует, связываясь с метилированным гистоном Н3
• Белок RAP1 инициирует образование гетерохроматина у дрожжей, связываясь со специфическими последовательностями ДНК
• Мишенями для RAP1 служат теломерные повторы и молчащие последовательности, расположенные в HML и HMR
• RAP1 участвует в отборе белков SIR3/SIR4, которые взаимодействуют с N-терминальными участками Н3 и Н4
Инактивация хроматина происходит вследствие добавления белков к фибрилле нуклеосом. Она может быть вызвана многими причинами, например конденсацией хроматина, в результате которой он становится недоступным для аппарата генной экспрессии, присоединением белков, непосредственно блокирующих доступ к регуляторным сайтам, или же непосредственно ингибирующих транскрипцию.
Известны две системы инактивации хроматина: НР1 у млекопитающих и комплекс SIR у дрожжей. Хотя белки, входящие в их состав, различаются, процесс осуществляется по общему механизму: точками контакта в хроматине служат N-терминальные участки молекул гистонов.
Белок НР1 (гетерохроматиновый белок 1) был идентифицирован в составе гетерохроматина при окрашивании политенных хромосом соответствующими антителами. У дрожжей 5. pombe гомолог этого белка кодируется геном swi6. Белок, первоначально идентифицированный как НР1, в настоящее время называется HP1а, поскольку в дальнейшем были обнаружены еще два близких белка, НР1b и НР1y.
Белок НР1 содержит область, которая называется хромодомен и располагается вблизи N-концевого участка молекулы, а также связанный с ней другой домен, который называется хромотеневым и располагается на карбоксильном участке молекулы. О важности хромодомена свидетельствует тот факт, что в нем локализовано большинство мутаций, нарушающих функционирование белка НР1. Хромодомен состоит из 60 аминокислот. Он обнаружен в белках, принимающих участие как в активации, так и в репрессии хроматина, что позволяет предполагать его роль в белок-белковых взаимодействиях. Хромодомен(ы) обеспечивают взаимодействие белков с гетерохроматином за счет узнавания метилированных остатков лизина в гистоновых «хвостах».
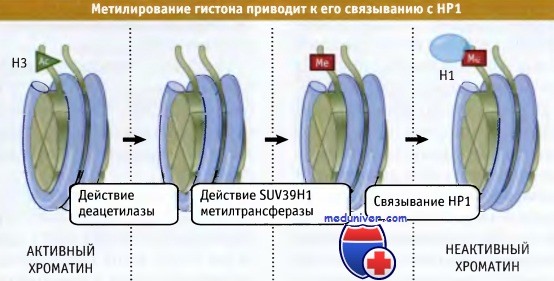
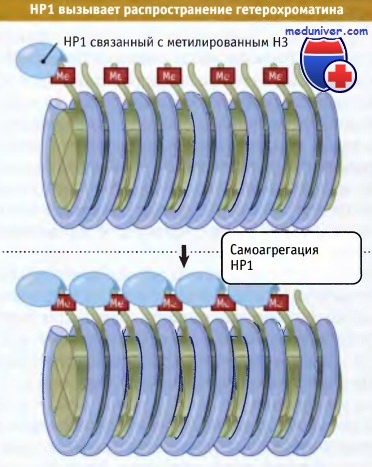
Мутации по ацетилазе, затрагивающие H3Ac-14Lys, предотвращают метилирование 9Lys. Гистон Н3, который метилируется по 9Lys, связывает белок НР1 через хромодомен. На рисунке ниже представлена модель образования гетерохроматина. Вначале происходит удаление ацетильной группы с 14Lys с помощью деацетилазы. Затем при участии метил азы в «хвосте» гистона Н3 создается метилированный сайт, к которому присоединяется белок НР1. На рисунке ниже показано, что между хромодоменом и метилированным лизином происходит взаимодействие, которое служит событием, запускающим процесс формирования неактивного хроматина. Как показано на рисунке ниже, в дальнейшем неактивная область может распространяться по хроматину, за счет способности все новых молекул НР1 к взаимодействию между собой.
Существование общего механизма формирования молчащих последовательностей у дрожжей подтверждается зависимостью их активации от общего набора генетических локусов. Мутации по любому из этих генов вызывают активацию двух молчащих локусов (HML и HMR), а также ослабляют степень инактивации генов, расположенных поблизости от теломерного гетерохроматина. Поэтому продукты этих генов ответственны за поддержание неактивного состояния обоих типов гетерохроматина.
Фермент SUV39H1 представляет собой гистон метилтрансферазу,
которая метилирует Lys-9 в гистоне Н3.
НР1 связывается с метилированным гистоном.
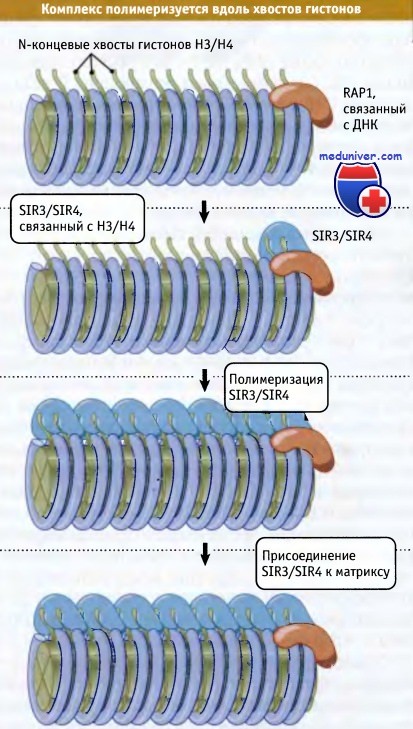
На рисунке ниже представлена модель, описывающая механизм действия этих белков. Лишь один из них способен связываться со специфической последовательностью ДНК. Это белок RAP1, который связывается с С1-3А повторами в теломерной области и присоединяется к цис-элементу сайленсинга, необходимому для подавления активности HML и HMR. Белки SIR3 и SIR4 реагируют с RAP1 и друг с другом (они могут функционировать как гетеромультимер). SIR3/SIR4 взаимодействуют с N-терминальными «хвостами» гистонов Н3 и Н4. (Фактически первое свидетельство в пользу непосредственного участия гистонов в формировании гетерохроматина было получено при исследовании мутаций, активирующих локусы HML и HMR, которые расположены в области генов, кодирующих синтез Н3 и Н4.)
Критическую роль в узнавании последовательностей ДНК, участвующих в формировании участков гетерохроматина, играет белок RAP1. Он обеспечивает взаимодействие белков SIR3/SIR4 с гистонами Н3 и Н4. Когда они связались с гистонами, образование комплекса может продолжаться и распространяться по хроматину дальше. Это может приводить к инактивации участка хроматина либо из-за ингибирующего эффекта SIR3/SIR4, либо из-за того, что связывание с гистонами НЗ/Н4 вызывает дальнейшие изменения в структуре хроматина. Мы не знаем, какие факторы ограничивают распространение комплекса. С-концевой участок молекулы SIR3 по составу близок к ядерным ламинам (компонентам ядерного матрикса) и может участвовать в фиксации гетерохроматина на периферии ядра.
Сказанное также справедливо для молчащих локусов HMR и HML. В образовании комплекса участвуют три фактора, проявляющих специфичность к определенным последовательностям ДНК: RAP1, ABF1 (фактор транскрипции) и ORC (комплекс начала репликации). В этом случае белок SIR1 взаимодействует с фактором, связанным с определенной последовательностью, и запускает процесс образования репрессорной структуры, состоящей из белков SIR2, SIR3 и SIR4. Белок SIR2 представляет собой деацетилазу гистонов. Реакция деацетилирования необходима для поддержания связывания белка SIR с хроматином.
Каким образом комплекс сайленсинга подавляет активность хроматина? Этот комплекс может вызвать такую конденсацию хроматина, что регуляторные белки окажутся не в состоянии найти свои мишени. Проще всего было бы предположить, что присутствие комплекса несовместимо с факторами транскрипции и РНК-полимеразой. Возможно, что комплексы блокируют ремоделирование хроматина (и таким образом предотвращают связывание этих факторов) или же непосредственно блокируют сайты связывания, тем самым предотвращая присоединение к ним факторов транскрипции.
Впрочем, все может быть не так просто, поскольку факторы транскрипции и РНК-полимеразу можно обнаружить в промоторной области неактивного хроматина. Это позволяет считать, что комплекс сайленсинга предотвращает функционирование факторов, а не их связывание с хроматином. Фактически может наблюдаться конкуренция между активаторами генов и репрессорами хроматина, за счет которой при активации промотора распространение комплекса сайленсинга по хроматину ингибируется.
Образование особой структуры хроматина на центромерных участках может быть связано с образованием гетерохроматина в определенной области. В клетках человека в области центромеры содержится специфический белок, CENP-B, необходимый для запуска модификации гистона Н3 (деацетилирование 9Lys и 14Lys, с последующим метилированием 9Lys), необходимой для связывания с белком Swi6, которое приводит к формированию гетерохроматина в данном участке.
В результате метилирования гистона Н3 на нем создается сайт связывания для НР1.
В результате связывания НР1 с метилированным Н3 гистоном образуется триггер,
способный запускать инактивацию генов, поскольку присоединение дополнительных молекул НР1 приводит к их агрегации на нуклеосомной цепи.
Связывание RAP1 с ДНК приводит к образованию гетерохроматина.
SIR3/SIR4 связывается с RAP1 и с гистонами Н3/Н4.
Комплекс полимеризуется вдоль хроматина и может связывать теломеры с ядерным матриксом.