Механизм передачи сигнала вторичными мессенджерами
• Вторичные мессенджеры распространяют сигналы между белками, которые находятся на определенном расстоянии друг от друга.
• ЦАМФ и Са2+ представляют собой распространенные вторичные мессенджеры.
При передаче сигналов используются как белки, так и небольшие молекулы, обладающие характерными свойствами. Небольшие молекулы, которые функционируют как внутриклеточный сигнал, или вторичный мессенджер, имеют ряд преимуществ перед белками в качестве сигнального интермедиата. Небольшие молекулы быстро образуются и столь же быстро распадаются.
Поскольку они синтезируются быстро, они могут действовать в высоких концентрациях, так что их сродство к белкам-мишеням может быть невысоким. Низкое сродство позволяет малым молекулам моментально диссоциировать так, что передача сигнала может быть быстро прервана, когда свободные молекулы вторичного мессенджера распадаются или инактивируются.
Поскольку молекулы вторичного мессенджера невелики, они способны быстро распространяться по клетке, хотя во многих клетках существуют механизмы, препятствующие этому. Таким образом, при выработке быстрой реакции на сигнал, особенно связанной с преодолением расстояния, вторичные мессенджеры обладают рядом преимуществ перед белками.
Вторичные мессенджеры используются также, когда необходимо одновременно адресовать сигналы многим белкам-мишеням. Эти их преимущества часто компенсируют отсутствие у них каталитической активности и неспособность одновременно связывать несколько молекул.
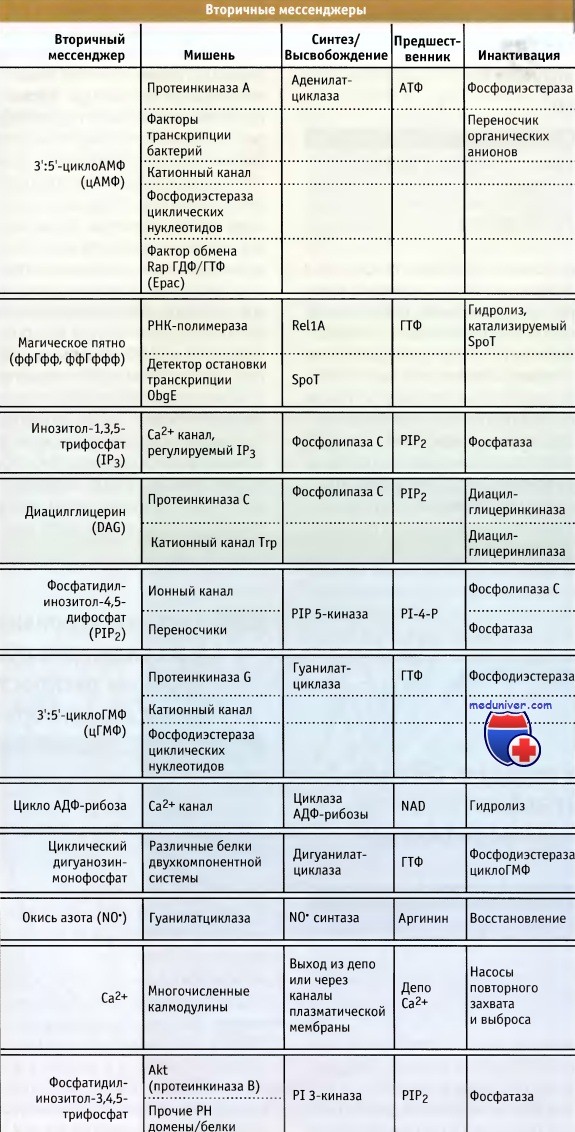
На рисунке ниже представлены внутриклеточные вторичные мессенджеры, возникшие в клетках в ходе эволюции. Количество их невелико, что вызывает удивление. Некоторые представляют собой нуклеотиды, образовавшиеся из основных метаболических предшественников. Это цАМФ, цГМФ, ффГффф и циклоАДФ-рибоза. Прочие растворимые вторичные мессенджеры включают фосфорилированный сахар, инозитол-1,4,5-трифосфат (IP3) и двухвалентный ион Са2+, а также свободный радикал, окись азота (NO).
К вторичным мессенджерам липидной природы относятся диацилглицерин и фосфатидилинозитол-3,4,5-трифосфат, фосфатидилинозитол-4,5-дифосфат, сфингозин-1-фосфат и фосфатидная кислота.
К числу первой из сигнальных молекул, описанной как вторичный мессенджер, относится цАМФ. Это название он получил потому, что цАМФ образуется в клетках животных как второй внутриклеточный сигнал, в ответ на действие многих внеклеточных гормонов, т. е. первых мессенджеров на пути передачи сигнала. цАМФ принимает участие в передаче информации различным регуляторным белкам у прокариот, грибов и в клетках млекопитающих (его присутствие у высших растений пока не доказано).
Активность аденилатциклазы, фермента который синтезирует цАМФ из АТФ, регулируется различными путями, в зависимости от организма, в котором она функционирует. У животных аденилатциклаза представляет собой интегральный белок плазматической мембраны, и ее множественные формы активируются при действии различных агентов. В клетках животных аденилатциклаза обычно активируется под действием Gs, который первоначально был описан как регулятор активности фермента. Некоторые ферменты грибов также активируются G-белками. Циклазы бактерий характеризуются более разнообразными системами регуляции активности.
цАМФ удаляется двумя способами. Он может выходить из клетки под действием АТФ-зависимого анионного насоса, но гораздо чаще гидролизуется представителями семейства фосфодиэстераз циклических нуклеотидов. Это большая группа белков, которые сами находятся под множественным регуляторным контролем.
Прототипом регулятора цАМФ у животных является цАМФ-зависимая протеинкиназа, однако вскоре после идентификации этого фермента у бактерий был обнаружен цАМФ-регулируемый фактор транскрипции, а в настоящее время известны и другие эффекторы. Система цАМФ остается прототипом сигнальной системы эукариот. Это выражается в том, что ее компоненты представляют собой почти все известные разновидности сигнальных молекул и демонстрируют все виды их взаимодействий: гормон, рецептор, G-белок, аденилатциклаза, протеинкиназа, фосфодиэстераза и выводящий насос.
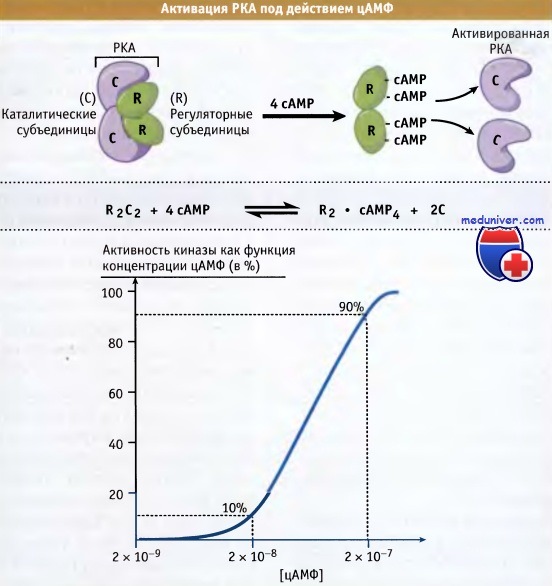
Протеинкиназа РКА, активность которой стимулируется вторичным мессенджером, представляет собой тетрамер, состоящий из двух каталитических (С) и двух регуляторных (R) субъединиц. Субъединица R связывается с каталитической субъединицей посредством сайта связывания субстрата, и, таким образом, С поддерживается в неактивном состоянии. Каждая субъединица R связывает две молекулы циклического нуклеотида, т. е. четыре молекулы цАМФ на холофермент РКА. Когда сайты связывания оказываются заполненными, димер субъединиц R быстро диссоциирует, оставляя две высокоактивные свободные каталитичнеские субъединицы.
Сродство R к С в присутствии и в отсутствие цАМФ, различается примерно в 10 000 раз. Выраженный кооперативный характер связывания цАМФ приводит к очень крутому подъему кривой активации и к порогу, ниже которого существенной активации РКА не происходит. Таким образом, активность РКА резко возрастает в очень узком диапазоне концентрации цАМФ. РКА также регулируется за счет фосфорилирования петли активации. Фосфорилирование происходит одновременно с трансляцией и необходимо для сборки тетрамера R2C2.
В основном РКА представляют собой цитозольные белки и занимают в клетке специфические места, связываясь с каркасными структурами органелл (белки, закрепляющие А-киназу, или АКАР). Эти АКАР способствуют фосфорилированию мембранных белков, включающих GPCR, переносчики и ионные каналы. Они также обеспечивают локализацию РКА в других местах: в митохондриях, цитоскелете и в центросомах. АКАР часто содержат сайты связывания для таких регуляторных молекул, как фосфопротеинфосфатазы и другие протеинкиназы, которые необходимы для координации функционирования нескольких сигнальных путей и интеграции выходных сигналов.
Обычно РКА фосфорилируют субстраты с участием последовательности Arg-Arg-Xaa-Ser-гидрофобный остаток, которая у большой группы киназ узнает остатки основных аминокислот, расположенные перед сайтом фосфорилирования. РКА участвует в регуляции активности различных внутриклеточных белков, от ионных каналов до факторов транскрипции, и благодаря субстратной специфичности фермента, часто, на основании анализа первичной структуры белка, можно предсказать, является ли он субстратом для киназы.
РКА фосфорилирует Ser 133 в CREB, представляющем собой белок, связывающийся с цАМФ-зависимым элементом, который в основном обеспечивает влияние цАМФ на транскрипцию многих генов.
Основные внутриклеточные вторичные мессенджеры, некоторые белки,
активность которых они регулируют, их источники и функционирование.
РКА представляет собой гетеротетрамер, который состоит из двух каталитических (С) и двух регуляторных (R) субъединиц.
Связывание четырех молекул цАМФ с регуляторными субъединицами приводит к диссоциации двух молекул С,
т. е. к активной форме РКА, образующейся из димера регуляторных субъединиц, связанных с цАМФ.
На нижнем рисунке показано, что кооперативное связывание четырех молекул цАМФ приводит к резкой активации киназы.
При увеличении концентрации цАМФ только в 10 раз, активность киназы возрастает с 10 до 90%.
Появление порога объясняется небольшими изменениями активности при низких концентрациях цАМФ.