• Для выхода клетки из митоза необходима инактивация Cdk1
• При выходе из митоза необходимо дефосфорилирование белков
• Инактивация Cdk1 и преодоление последствий фофсфорилирования белков скоординированы с разборкой митотического веретена и цитокинезом
Мы рассмотрели основные молекулярные механизмы и изменения в клеточных структурах, связанные с наступлением митоза. Для завершения цикла, клетка должна выйти из митоза и вернуться в интерфазу. Сейчас мы расскажем, как это происходит.
Для митоза характерна высокая активность Cdk1. Чтобы клетки вернулись в интерфазу и вступили в новый цикл, должна произойти инактивация Cdk1 и дефосфорилирование белков. Этот процесс возвращения в интерфазу называется выходом из митоза. Как отмечалось ранее, инактивация Cdk1 происходит в основном за счет убиквитин-зависимого протеолиза циклина В. Впрочем, в этом процессе также участвуют и другие механизмы.
Дополнительный путь преодоления последствий активации Cdk1 был обнаружен на мутанте S. cerevisiae, cdc14-3, рост которого остановлен в телофазе, но клетки обладают высоким уровнем активности Cdk1. Было показано, что cdc14 содержит фосфатазу, обладающую двойной специфичностью (способную дефосфорилировать остатки серина, треонина и тирозина), которая дефосфорилирует субстраты Cdk1. Среди субстратов этой фосфатазы находится фактор транскрипции Swi5, активатор АРС, Cdh1 и ингибитор Cdk1 (CKI), Sic1.
После дефосфорилирования Swi5 накапливается в ядре, в то время как фосфорилирование Cdk1 ингибирует его поступление в ядро. Находясь в ядре, Swi5 увеличивает экспрессию Sic1. Вместе с тем, дефосфорилирование Sic1 под действием Cdc14 предотвращает его деградацию SCF. Таким образом, Cdc14 и Swi5 действуют согласованным образом, ингибируя Cdk1 за счет увеличения уровня одного из ее ингибиторов.
Это приводит к тому, что даже если циклин неполностью деградирует под действием АРС, то все равно происходит инактивация Cdk1-циклинового комплекса. Вместе с тем, дефосфорилирование субстрат-специфического активатора АРС, Cdh1, под действием Cdc14, позволяет ему связаться с АРС и пометить митотические циклины для протеолиза. Действуя таким образом, АРС продолжает активироваться по мере снижения уровня Cdk1, и активность Cdc14 увеличивается.
Последние исследования гомологов Cdc14 в клетках эукариот показывают, что способность этого семейства фосфатаз к дефосфорилировнию белков сохраняется. Однако в большинстве исследованных клеток эта фосфатаза не является единственной. Поэтому, в преодоление последствий активации Cdkl, а также других киназ митоза, таких как Plk, NEK и Aurora, важную роль должны играть другие фосфатазы.
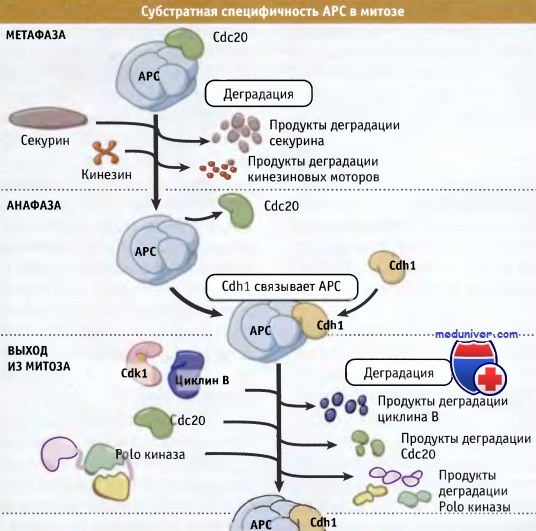
АРС Cdc20 обеспечивает протеолиз секурина, ряда белковых моторов, относящихся к группе кинезина, и небольшого количества циклина В.
В течение большей части клеточного цикла, включая ранние фазы митоза, еще один активатор АРС, Cdh1 фосфорилируется Cdk1 и может связываться с АРС.
Снижение активности Cdkl в анафазе, обусловленное инициацией протеолиза циклина В под действием АРС Cdc20,
и активация фосфатазы Cdc14 приводят к дефосфорилированию Cdh1.
Затем этот дефосфорилированный белок связывается с АРС, и АРС Cdh1 обеспечивает протеолиз циклина и инактивацию Cdk1.
Протеинфосфатаза 2А (РР2А), наряду с выполнением других функций, участвует в регуляции клеточного цикла. Дрожжевые мутанты, с нарушениями функции этого фермента, обнаруживают разнообразные дефекты клеточного цикла. Эта очень распространенная фосфатаза обладает несколькими регуляторными субъединицами и действует на многие субстраты. Однако наличие большого количества регуляторных субъединиц и влияние РР2А на множество клеточных процессов затрудняет понимание ее роли в процессах, приводящих клетку к выходу из митоза.
Аналогичным образом, непростую задачу представляет собой выяснение роли в митозе протеинфосфатазы I (PPI), у которой имеется еще больше регуляторных субъединиц, чем у РР2А. Тем не менее генетические и биохимические исследования подтвердили ее участие в регуляции митоза. PPI участвует в дефосфорилировании гистона Н3, фосфорилированного с участием киназы Aurora, и в инактивации этой киназы.
Действительно, на почкующихся дрожжах было показано, что PPI и киназа Aurora взаимодействуют друг с другом, и что некоторые процессы митоза регулируются балансом активности двух ферментов. Однако, по-видимому, фосфорилирование осуществляется не только за счет киназы Aurora, и при выходе клетки из митоза дефосфорилирование соответствующих субстратов также происходит с участием PPI.
Интересно, каким образом скоординировано разделение сестринских хроматид с инактивацией Cdk1. Этот вопрос имеет принципиальное значение, поскольку инактивация Cdk1 до разделения сестринских хроматид привела бы к ошибочной сегрегации хромосом за счет преждевременной разборки веретена и наступления цитокинеза. Один из предполагаемых механизмов состоит в постепенной деградации белков, которые являются субстратами АРС.
Например, в почкующихся дрожжах деградация секурина и циклина В происходит в разное время и при действии различных форм АРС (протеолиз секурина происходит под действием APCcdc20, а основное количество циклина В разрушается APCcdh1). В исследованиях на почкующихся дрожжах также показано, что стабильный секурин ингибирует протеолиз циклина. Таким образом, у дрожжей, деструкция секурина под действием APCcdc20 приводит к разделению сестринских хроматид и к снятию ингибирования протеолиза циклина.
Вместе с тем, поскольку основное количество циклина при переходе от метафазы к анафазе оказывается стабильным, Cdk1 находится в активном состоянии, и Cdhl поддерживается в фосфорилированной форме, что предотвращает его взаимодействие с АРС. Продолжающееся фосфорилирование Cdh1 до начала снижения активности Cdk1, и активация Cdcl4 гарантируют, что основная масса протеолиза циклина, и отсюда инактивация Cdk1, происходит только после деградации секурина и расхождения хромосом. Порядок наступления событий протеолиза и субстраты АРС в клетках почкующихся дрожжей представлены на рисунке ниже.
Не известно, насколько эта схема приложима ко всем организмам, однако она наглядно иллюстрирует всю сложность процессов, участвующих в регуляции ключевых процессов клеточного цикла.
Как секурин влияет на деградацию циклина в клетках почкующихся дрожжей? Как отмечалось ранее, при деградации секурина высвобождается активная форма сепаразы. Сепараза расщепляет когезин, однако, по крайней мере у почкующихся дрожжей в ранней анафазе, она еще запускает выход из ядрышек Cdc14 фосфатазы. Выход Cdc14 из ядрышек приводит к ее активации. После этого Cdc14 распределяется по клетке и поддерживается в активной форме с помощью сигнального каскада, который называется сетью выхода из митоза (англ. mitotic exit network, MEN). Поэтому при активации сепаразы запускаются все процессы, приводящие клетку к выходу из митоза.
Сигнальные процессы MEN участвуют в цитокинезе и в выходе клетки из митоза. Аналогичные процессы контролируют наступление цитокинеза и образование перегородки у сливающихся дрожжей. Эти процессы называются сеть инициации образования перегородки (англ. septation mitiation network, SIN). Происходящие процессы представляют собой каскад протеинкиназных сигналов, который регулируется небольшой ГТФазой. Общая цель MEN и SIN, локализованная на полюсах веретена, позволяет скоординировать цитокинез с сегрегацией хромосом.
Учитывая необходимость такой координации, вполне вероятно. что аналогичные процессы происходят у многоклеточных эукариот. Действительно, в клетках растений и животных были обнаружены гомологи некоторых из компонентов сетей. Процессы, участвующие в сети MEN и SIN, представлены на рисунке ниже. Одним из важнейших остается вопрос о специфических мишенях процессов MEN и SIN, необходимых для запуска цитокинеза.
Суммируя вышеизложенное, отметим, что критическим для клетки событием является инактивация Cdk1 и дефосфорилировние субстратов, с тем чтобы разделение сестринских хроматид, разборка веретена и цитокинез проходили в скоординированном режиме.
Первый видеокадр, показывающий хромосомы в начальных стадиях митоза.