Отклик неструктурированных белков (unfolded protein response, UPR) - реакция на несвернутые белки
• Образование нативной структуры в люмене ЭПР контролируется особым процессом, получившим названия «отклик неструктурированных белков (UPR)» или «реакция на несвернутые белки». При этом запускается каскад процессов, приводящих к увеличению экспрессии генов шаперонов ЭПР.
• Белок Ire1p опосредует отклик неструктурированных белков у дрожжей и активируется в ответ на стрессорные условия.
• Активированный Ire1p осуществляет сплайсинг иРНК НАС1, что приводит к образованию Had белка, фактора транскрипции, локализованного в ядре и связанного с промотором генов, содержащих UPR повтор.
• Отклик неструктурированных белков у высших эукариот включают больше уровней, чем для клеток дрожжей.
Даже при функционировании системы ERAD в эндоплазматическом ретикулуме (ЭПР) могут накапливаться белки, обладающие неправильной нативной структурой. Это происходит в случаях, когда клетка испытывает стресс, например при вирусной инфекции, повышенной температуре или при воздействии некоторых химических веществ. При этом утрачиваются механизмы контроля за конформацией белка и нарушаются процессы секреции. Поскольку белки секреторного пути должны попасть в ЭПР, эта органелла играет важную роль в поддержании гомеостаза на всем секреторном пути. ЭПР узнает нарушения процесса секреции белков, которые происходят в различных участках секреторного пути. Отклик неструктурированных белков (англ. unfolded protein response, UPR) представляет собой каскад процессов, происходящих на пути от люмена ЭПР до ядра клетки, в результате которого в ЭПР поддерживается контроль над условиями созревания белков и при необходимости усиливается экспрессия шаперонов. Существование этого отклика характерно для всех клеток эукариот, однако наиболее существенные его детали на молекулярном уровне впервые были выяснены на дрожжевых клетках.
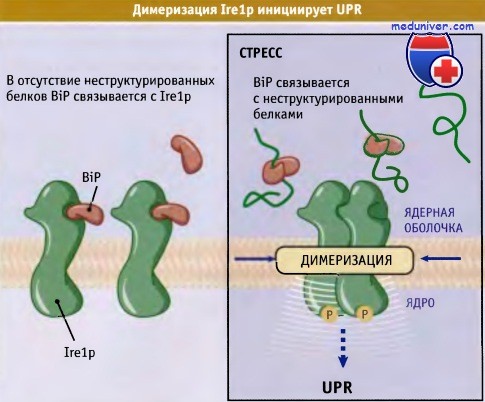
Для UPR дрожжей критическим медиатором сигнала, т. е. продуктом, который «отслеживает» условия для сворачивания белков в ЭПР и передает эту информацию, является белок мембраны ЭПР, называемый Ire1p. Этот белок способен образовывать димеры за счет ассоциации со своими доменами со стороны люмена, однако в нормальных условиях с этими доменами связывается BiP, что делает невозможным образование димеров. Однако при стрессе BiP связывается с неправильно скрученными белками, a Ire1p образует димеры и тем самым активирует каскад процессов, составляющих UPR. Сигнал передается через цитозольный домен Irelp, который представляет собой серин-треонин киназу. (Возможно, что Ire1p расположен в области ЭПР, образующей ядерную оболочку, и поэтому его «цитозольный» домен в действительности находится в ядре, хотя это строго и не доказано.)
Димеризация Ire1p индуцирует процесс аутофосфорилирования и активации цитоплазматического домена. Поэтому Ire1p функционирует, как и другие рецепторные киназы, за исключением того, что димеризуется в отсутствии, а не в присутствии лиганда (BiP). Он также передает сигнал между компартментами, а не с поверхности внутрь клетки.
В обычных условиях BiP может связаться с Ire1p и тем самым предотвратить его димеризацию.
При развитии стресса BiP связывается с неструктурированными белками.
Это приводит к димеризации Irel1p и к развитию отклика неструктурированных белков.
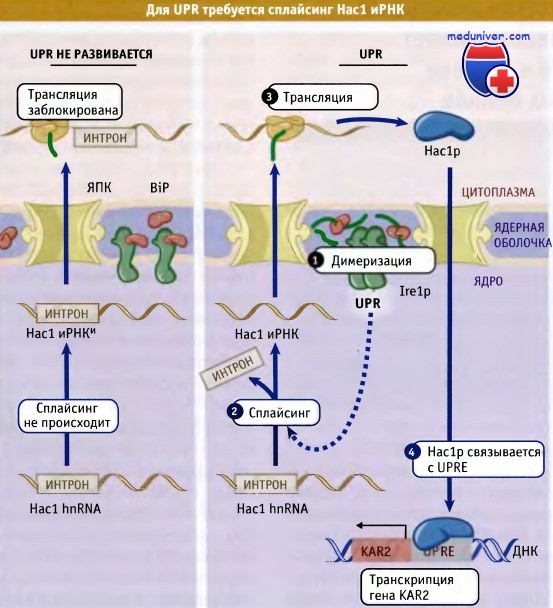
Как показано на рисунке ниже, при активации Ire1p второй домен цитозольной области этого белка катализирует удаление интрона из иРНК специфического гена, НАС1. Когда интрон удален, лигаза тРНК соединяет эк-зоны, образуя новую иРНК. Таким образом, сплайсинг с участием Ire1p отличается от сплайсинга, катализируемого сплайсосомой. Сплайсинг НАС1 иРНК является критическим событием для UPR, поскольку интрон задерживает рибосому на иРНК и нарушает трансляцию белка Нас1р. Однако после удаления интрона этот белок транслируется.
Белок Hac1p представляет собой фактор транскрипции, который связывается с промотором гена, кодирующего белок BiP, и стимулирует его транскрипцию. (Дрожжевой гомолог BiP называется KAR2.) Связывание происходит с участием специфической регуляторной последовательности, которая носит название элемента отклика неструктурированных белков (англ. unfolded protein response elements, UPRE). Эта последовательность также присутствует в промоторной области некоторых других генов, и Hac1p также стимулирует транскрипцию этих генов. Таким образом, узнавая неправильно структурированные белки посредством их связывания с BiP, Ire1p запускает каскад событий, которые приводят к дополнительному синтезу ЭПР шаперонов.
Эти шапероны усиливают способность клеточных контрольных систем справляться с притоком большого количества неструктурированных белков, при попадании клетки в условия стресса. Ответная реакция клетки носит более обширный характер, чем было выше описано. Хотя первоначально существование UPR было обнаружено по его влиянию на экспрессию специфических шаперонов, сейчас известно, что этот отклик затрагивает экспрессию сотен различных генов. Исследование природы экспрессируемых генов позволяет предполагать, что при появлении неструктурированных белков в клетке меняются элементы трансляции, транслокации, секреции и пролиферации мембран ЭПР. К числу такого рода ответов относится индукция синтеза липидов, необходимая для увеличения массы мембран ЭПР.
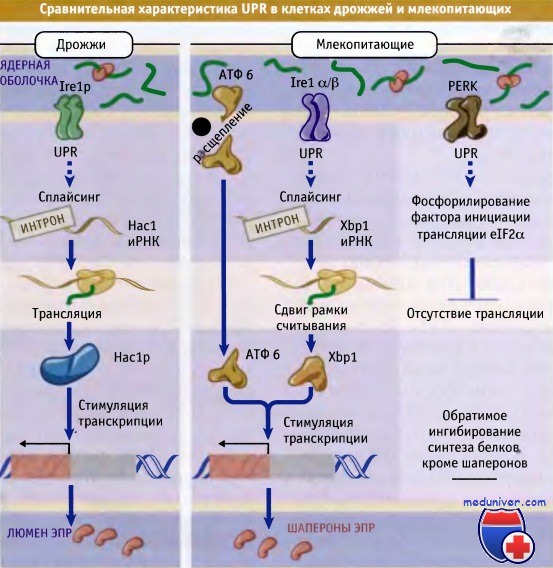
UPR в клетках млекопитающих похож на UPR клеток дрожжей, однако имеет более сложный характер. Гомологи Ire1p у млекопитающих также катализируют сплайсинг, который приводит к увеличению экспрессии шаперонов ЭПР, однако при этом транскрипционный фактор отличается от Hac1p. Этот фактор в клетках млекопитающих называется XbpI. Однако белок IRE1, по-видимому, не играет критической роли в UPR клеток млекопитающих, поскольку клетки, дефицитные по этому белку, не утрачивают способность к активации UPR в условиях стресса и стимулируют синтез шаперонов. Согласно современному представлению, IRE1 у млекопитающих контролирует экспрессию генов, необходимых для стимуляции ERAD. Вероятно, белок IRE1 и его мишень ХВР1 играют важную роль в созревании В-клеток, у которых масса ЭПР должна значительно увеличиваться по мере приобретения способности к синтезу и секреции антител.
В UPR у млекопитающих участвуют нехарактерные для дрожжевых клеток компоненты и процессы. При стрессе в ЭПР активируется второй фактор транскрипции, который носит название ATF6. Обычно этот белок пронизывает мембрану ЭПР, однако индукция UPR приводит к отщеплению его мембранного домена. Высвободившийся при этом цитозольный домен транспортируется в ядро, где связывается с промоторами генов, экспрессирующих шапероны ЭПР. Наряду с усилением контроля качества при ЭПР-стрессе, в клетках млекопитающих снижается синтез белка, и в ЭПР начинает поступать меньше белков. В этом процессе принимает участие белок PERK, представляющий собой трансмембранную киназу ЭПР, домен которой со стороны люмена гомологичен белку Irelp, но цитозольный домен имеет отличную структуру.
PERK в повышенных количествах экспрессируется в клетках поджелудочной железы, и его инактивация приводит к дисфункции органа. Это свидетельствует о значимости UPR для органов, характеризующихся высокой секреторной активностью. Активированный цитозольный домен белка PERK фосфорилирует а-субъединицу фактора инициации трансляции eIF2. Это приводит к общему ингибированию синтеза белка, в результате чего уменьшается его количество, поступающее в ЭПР. Важно, что этот эффект носит обратимый характер, и, таким образом, увеличение синтеза шаперонов носит регулируемый характер. Вместе с тем, по крайней мере одна иРНК, кодирующая фактор транскрипции ATF4, может транслироваться только при фосфорилировании eIF2a и ингибировании общего синтеза белка. ATF4, который синтезируется в результате фосфорилирования eIF2a, транспортируется в ядро и регулирует экспрессию генов, участвующих в энергетических процессах, поддержании окислительно-восстановительного потенциала и синтезе аминокислот.
Следует отметить, что при иных типах стресса другие киназы фосфорилируют eIF2a (например, при вирусной инфекции активируется для фосфорилирования eIF2a киназа PKR), и, таким образом, мишенями для ATF4 служат те гены, которые важны для общей реакции клетки на стресс, в то время как пути через ATF6 и IRE1 представляются более ЭПР-стресс-специфичными.
С активацией процессов UPR связано развитие многих патологических состояний, включающих нейродегенеративные болезни, диабет, функциональные расстройства печени, нарушения свертываемости крови, и многие другие. В экспериментальных условиях постоянная активация UPR приводит к активации процесса клеточной гибели. Однако развитие большинства болезней, связанных с нарушением UPR, происходит через несколько месяцев или лет. Это позволяет предполагать, что физиологическая активация UPR скорее напоминает адаптивный ответ, чем апоптоз. Учитывая, что UPR активирует как процессы выживаемости, так и апоптоз, не вполне ясно, каким образом этот процесс может способствовать выживаемости клетки в условиях хронического стресса.
Димеризация Ire1p вызывает сплайсинг Had иРНК и ее трансляцию.
Образуется фактор транскрипции Hac1p, который стимулирует экспрессию генов, кодирующих шапероны ЭПР.
У высших эукариот существует несколько путей UPR.
Они включают ингибирование трансляции через PERK и расщепление ATF6 с образованием фактора транскрипции.