• Убиквитин и родственные ему белки могут ковалентно присоединяться к другим белкам и служить метками мишени
• Убиквитин узнается различными белками, которые с ним связываются
• Убиквитинилирование происходит наряду с другими ковалентными модификациями
• Наряду с обоспечением деградации белков, их убиквитинилирование играет роль в системах передачи сигналов
Важнейшим механизмом контроля функций белков является их ковалентная модификация с помощью небольших белков, относящихся к семейству убиквитина. Убиквитин представляет собой одно из семейств белков, которые носят название убиквитин-подобные белки (Ubl).
Сам убиквитин обладает весьма консервативной структурой и обнаружен во многих объектах, что свидетельствует о функциональной значимости всех 76 остатков аминокислот, из которых он состоит. Наряду с давно известной ролью убиквитина в инициации деградации белков, он также обладает многочисленными функциями в системе передачи сигнала.
Ubl белки образуют конъюгат с субстратным белком за счет образования изопептидной связи между аминогруппой субстрата и С-концевым остатком Gly белка Ubl. Образование конъюгата катализируется белками E1, Е2 и Е3. К одному субстрату могут присоединиться несколько Ubl белков, часто при этом образуются полиубиквитиновые цепи. Моно- и полиубиквитинилирование изменяют свойства белка и индуцируют сигналы, распространяющиеся далее.
Моноубиквитинилирование представляет собой регуляторную модификацию, которая играет важную роль в везикулярном переносе и репарации ДНК. Например, моноубиквитини-лированная форма белка FANCD2 ассоциирует с белком репарации BRCA1 на сайтах, где происходит репарация ДНК. Модификация белком Ubl, SUMO, играет важную роль в ядерном транспорте, транскрипции и в прохождении клетки по циклу.
Образование цепей полиубиквитина происходит, когда убиквитинилированию подвергаются остатки Lys (особенно К48 и К63) в самом убиквитине. Формирование полиубиквитина на К48 обычно приводит к тому, что белок направляется в протеосомы для деградации, а конъюгация с полиубиквитиновыми остатками на К63 способствует передаче сигнала и не приводит к протеолизу. Связанный с белком убиквитин узнается различными убиквитин-связывающими доменами, включая UIM (последовательность, взаимодействующая с убиквитином), UBA (сайт ассоциации с убиквитином) и некоторыми доменами цинковых пальцев. Эти домены в модифицированных белках способны служить рецепторами для убиквитина.
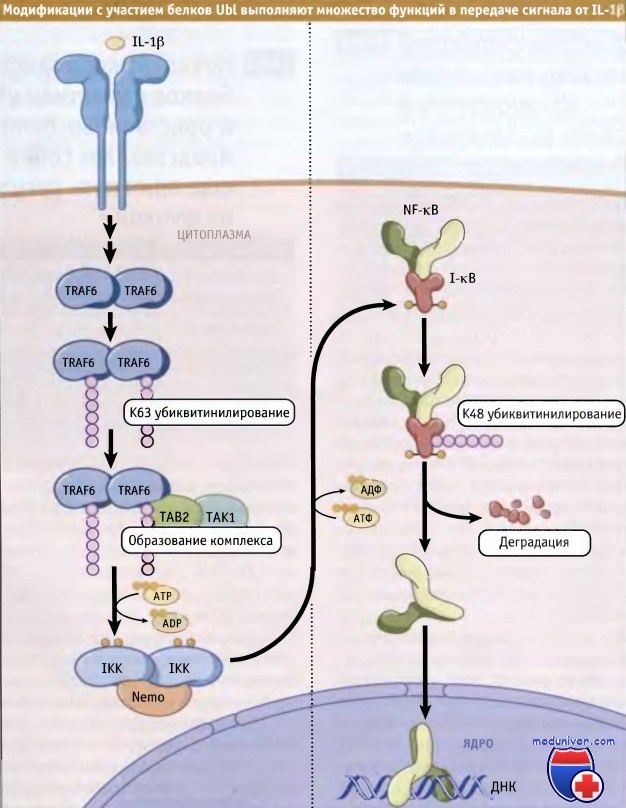
Активация фактора транскрипции NF-кВ происходит по механизму, связанному с модификацией, как за счет добавления белков Ubl, так и фосфорилирования. На рисунке ниже представлен пример регуляции активности этого фактора с участием убиквитина. До стимуляции NF-кВ удерживается в цитоплазме в неактивном состоянии за счет связывания со своим ингибитором, 1кВ. Фосфорилирование ингибитора 1кВ киназным комплексом (IKK) способствует его узнаванию мультисубъединичной Е3 лигазой, которая обеспечивает его убиквитинилирование и последующую деградацию в протеосомах. Деградация 1кВ позволяет NF-кВ мигрировать в ядро и участвовать в контроле транскрипции.
Стабилизация 1кВ происходит в ответ на определенные сигналы, за счет ковалентного присоединения SUMO, одного из белков Ubl. Этот белок присоединяется к тому же остатку Lys, который конъюгируется с убиквитином при подготовке 1кВ к деградации. Таким образом, SUMO стабилизирует 1кВ и ослабляет эффект NF-кВ. Это один из многочисленных примеров помех, возникающих при функционировании конъюгатов Ubl.
Ключевым регуляторным событием в передаче сигнала с участием NF-кВ является активация комплекса IKK. Этот комплекс сам регулируется за счет убиквитинилирования и фосфорилирования. Один из цитокинов, интерлейкин-1b (IL-1b), обеспечивает ассоциацию адаптерных белков со своим рецептором, образуя комплекс активации рецептора. Комплекс активации рецептора интерлейкин-1b мобилизует другой адаптерный комплекс, содержащий TRAF6. В результате фосфорилирования комплекс TRAF6 диссоциирует от комплекса активации рецептора и высвобождается в цитоплазму.
TRAF6 содержит домен RING и Е3 убиквитин лигазу, которая катализирует образование полиубиквитиновых цепей на К63 в молекуле протеинкиназы ТАК1. После этого, ТАК1 мобилизует ТАВ2 и ТАВ3, которые представляют собой адаптерные белки, содержащие консервативные домены цинковых пальцев. Эти домены связываются с плиубиквитинилированной молекулой ТАК1 и стимулируют ее активность. Активированная молекула ТАК1 фосфорилирует и активирует IKK, которая затем фосфорилирует 1кВ, подготавливая его к деградации.
Таким образом, такие домены связывания убиквитина, как цинковые пальцы в ТАВ2 и ТАВЗ могут избирательно узнавать полиубиквитиновые цепи на остатке К63 и способствовать передаче сигнала.
Активность убиквитин лигазы может непосредственно контролироваться небольшими природными молекулами. Ауксин (индолил-3-уксусная кислота) является растительным гормоном и регулирует процессы развития, способствуя транскрипции множества генов. Однако, ауксин не стимулирует активность факторов транскрипции, а скорее интенсифицирует деградацию некоторых ее специфических репрессоров. Фактически, рецепторы ауксина представляют собой убиквитин лигазный комплекс, который подготавливает к протеолизу репрессоры транскрипции, находящиеся под контролем ауксина. В экстрактах растительных клеток белки, связывающиеся с ауксином, относятся к белкам, содержащим домен F.
Активация NF-кВ включает этапы, зависящие от взаимодействия белков, связанных с убиквитином через убиквитин-связывающие белки,
конкуренции с присоединением SUM0, фосфорилирования и убиквитин-зависимой деградации белка.