Трансмембранный градиент ионов натрия и натрий зависимые переносчики
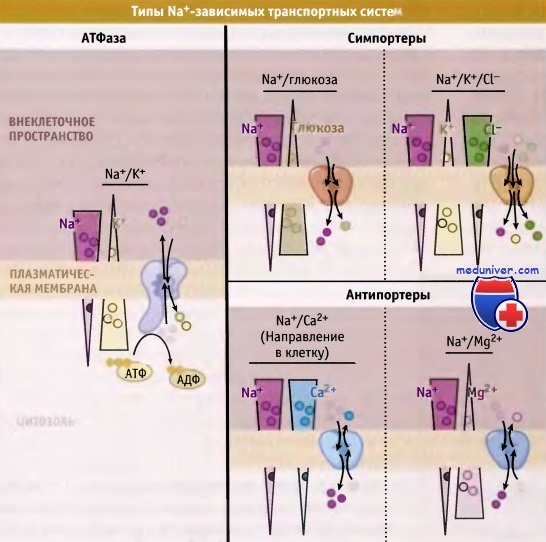
• Градиент ионов Na+ через плазматическую мембрану поддерживается за счет Na+/K+-АТФазы
• Энергия, которая высвобождается при транспорте Na+ в направлении градиента, расходуется на транспорт различных субстратов
• В желудочно-кишечном тракте усвоение сахара происходит за счет переносчика Na+/глюкозы
• Удаление ионов Са2+ из цитозоля возбудимых клеток осуществляется в основном посредством транспортного механизма с участием Na+/Са2+-обменника
• Внутриклеточная концентрация Cl- регулируется с помощью Na+/K+/Cl--котранспортеров
• Ионы магния транспортируются из клетки с помощью Na+/Mg2+-обменников
Клетки поддерживают трансмембранный градиент ионов Na+, направленный со стороны внеклеточного пространства. Существование такого градиента необходимо для протекания многих процессов мембранного транспорта, зависимых от Na+.
Электрохимический градиент Na+ на плазматической мембране поддерживается Na+/К+-АТФазой, которая представляет собой наиболее активный транспортный белок. Этот белок расходует энергию гидролиза АТФ на транспорт трех ионов Na+ из клетки в обмен на поступление туда двух ионов К+. Оба иона перемещаются против своих градиентов концентрации.
Примеры Na+-зависимых переносчиков.
В этих и во многих других транспортных системах используется энергия градиента Na+ по сторонам плазматической мембраны, который поддерживается Na+/K+-АТФазой.
Показано направление ионного градиента в типичной животной клетке.
Поскольку при этом суммарный положительный заряд клетки снижается, Na+ насос рассматривается как генератор энергии. Благодаря Na+/К+-АТФазе по обеим сторонам мембраны возникает разность потенциалов, причем внутренняя среда клетки, по сравнению с внешней, заряжена более отрицательно. Разница в зарядах составляет отрицательный мембранный потенциал покоя.
Иными словами, энергия, необходимая для поддержания электрохимического градиента Na+, запасается на плазматической мембране.
Существование электрохимического градиента ионов Na+ важно для поддержания физиологических функций клеток многих тканей. Некоторые из Na+ или потенциал-зависимых каналов и вторичных транспортных белков представлены на рисунке ниже. Для транспорта веществ против концентрационного градиента или для генерации электрических сигналов в форме потенциала действия они используют энергию, которая запасена в виде электрохимического градиента Na+, направленного внутрь клетки.
Две основные группы Na+-зависимых мембранных белков включают потенциал-зависимые Na+-каналы и эпителиальные Na+-каналы. К третьей группе относятся Na+/субстратные переносчики, которые в зависимости от степени сходства первичной структуры подразделяются на несколько семейств. В данном разделе мы рассмотрим некоторые из этих переносчиков. Na+-зависимые переносчики, участвующие в регуляции pH, рассматриваются в следующем разделе.
На рисунке ниже представлены некоторые Na+-зависимые переносчики субстратов. С эволюционной точки зрения большинство этих переносчиков не связаны между собой и поэтому различаются по природе переносимого субстрата или по механизму транспортного процесса. Na+/субстратные переносчики могут функционировать в процессах катаболизма, обеспечивая субстратами метаболические процессы в клетке.
Они используют Na+-градиент для транспорта в клетку или из нее таких субстратов, как ионы, сахара, аминокислоты, витамины и мочевина. К таким переносчикам относятся котранспортеры Na+/глюкоза и Na+/иодид. У некоторых бактерий в осмотической регуляции участвует котранспортер Na+/пролин. У других выживаемость при инфекции обеспечивается функционированием Na+/субстрат симпортеров.
Некоторые клетки, например клетки ворсинок кишечника, при всасывании используют трансмембранный градиент Na+ для транспорта сахаров против градиента концентрации. Котранспортер Na+/глюкоза обеспечивает усвоение из кишечника двух сахаров, D-глюкозы и D-галактозы. Этот котранспортер кодируется геном, относящимся к семейству SGLT1. Белки этого семейства имеют общую коровую структуру, состоящую из 13 трансмембранных спиралей.
Сайт связывания сахара и транспортный путь котранспортера Na+/глюкоза образованы четырьмя трансмембранными спиралями, поблизости от С-терминального участка. Предполагается, что связывание Na+ обеспечивается N-терминальным участком котранспортера Na+/глюкоза и что совместный транспорт Na+ и глюкозы происходит при взаимодействии N- и С-терминальных доменов белка. Основной вопрос заключается в том, каким образом перенос сахара сопряжен с электрохимическим градиентом Na+.
Современная модель предполагает, что внеклеточный ион Na+ связывается с незанятым сайтом на транспортере и вызывает изменения конформации, в результате которых происходит связывание сахара. При связывании сахара конформация изменяется еще раз, в результате чего Na+ и сахар оказываются на внутриклеточной стороне мембраны. После высвобождения Na+ и сахара в клетку пустой контранспортер Na+/глюкоза меняет свою конформацию, возвращаясь к исходной. В общем, конформационные изменения, постулируемые в этой модели, напоминают изменения в унипортере GLUT-1, который функционирует как переносчик глюкозы.
Многие внеклеточные сигналы индуцируют высвобождение внутриклеточного Са2+. В цитозоле покоящейся клетки поддерживается низкая концентрация ионов Са2+, порядка 10-7 М. При Са2+-зависимой передаче сигнала происходит активация, при которой концентрация этого катиона в цитозоле увеличивается за счет поступления извне и высвобождения из депо саркоплазматического ретикулума. Когда сигнал проходит, то Са2+ транспортируется из цитозоля через плазматическую мембрану и в саркоплазматический ретикулум.
К числу двух главнейших белков плазматической мембраны, удаляющих Са2+ в большинстве клеток животных, относится Na+/Са2+-обменник (NCX) и АТФ-зависимый Са2+-насос. Na+/Са2+-обменник обладает примерно в 10 раз более низким сродством к ионам Са2+, но характеризуется в 10-50 раз более высоким числом оборотов, чем Са2+ АТФаза плазматической мембраны. Поэтому в возбудимых клетках Na+/Ca2+-обменник представляет собой основную систему выведения Са2+ на внешнюю сторону плазматической мембраны.
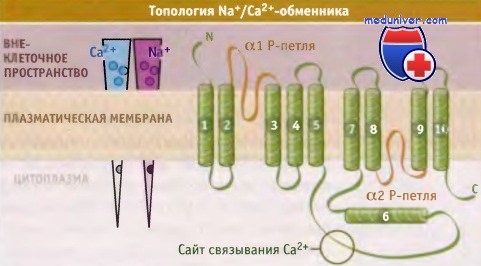
Предположительное строение Na+/Са2+-обменника клетки сердечной мышцы.
Показано направление градиентов Na+/Ca2+ в покоящейся животной клетке млекопитающих.
Na+/Са2+-обменники образуют семейство белков. Предполагается, что в клетках миокарда они состоят из девяти трансмембранных сегментов, большой внутриклеточной петли, принимающей участие в регуляции обмена, и антипараллельных доменов, содержащих a-повторы, которые необходимы для транслокации ионов. Две области а-повторов содержат участки трансмембранных сегментов 2, 3, 8 и 9 и, подобно аквапориновым водным каналам, имеют противоположную ориентацию.
Также обменник обладает двумя Р-петлями, содержащими Gly-Ile-Gly последовательности, напоминающие Gly-Tyr-Gly последовательности селективного фильтра К+-канала. Такая топологическая модель Na+/Са2+-обменника была предложена с учетом данных секвенирования, мутагенеза и опытов по функциональной активности. Таким образом, в отличие от переносчика глюкозы GLUT-1 и котранспортера Na+/глюкоза, предполагают, что Na+/Са2+-обменник имеет общие черты строения с канальными белками.
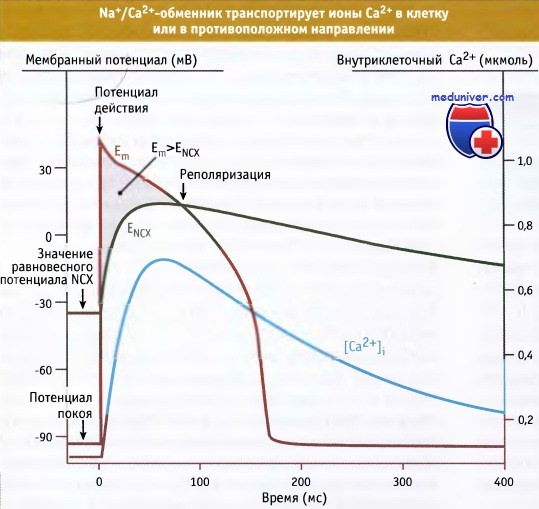
Считается, что Na+/Са2+-обменник транспортирует три иона Na+ в обмен на один ион Са2+, в результате чего на транспортный цикл возникает один положительный электрический заряд. Обменник может транспортировать ионы Са2+ или в клетку, или из нее, в зависимости от электрохимической движущей силы. Например, в клетках миокарда, находящихся в состоянии покоя, значение мембранного потенциала Ет меньше, чем значение равновесного потенциала для ЕNCX, и ионы Са2+ вытесняются из клетки.
Однако при генерации потенциала действия в фазе 0 и 1, Еm превышает ENCX, что вызывает кратковременный вход в клетку Са2+ через обменник. По мере реполяризации мембраны, мембранный потенциал опять принимает более отрицательное значение. (Em < ENCX), и ионы Са2+ снова выходят из клетки. Таким образом, ионы Са2+ на короткое время, перемещаемые обменником, меняют направление движения. Это происходит, когда мембранный потенциал принимает более положительное значение по отношению к обратному потенциалу Na+/Са2+-обменника.
Через несколько миллисекунд, в начале развития относительно длительного потенциала действия миокарда, при изменении трансмембранного Na+- или Са2+-градиента в клетки входят ионы Са2+. При физиологических условиях обменник в основном выполняет функцию вытеснения Са2+ из клетки за счет Na+-градиента. Однако положительные значения мембранных потенциалов при выходе на плато потенциала действия могут ограничивать выход Са2+.
Еще одним переносчиком, использующим энергию Na+-градиента для переноса ионов через плазматическую мембрану, является котранспортер Na+/K+/Cl-. Он осуществляет электронейтральный перенос ионов в стехиометрическом отношении 1Na+:1K+:2Cl-. Этот котранспортер катализирует поступление ионов в клетку в физиологических условиях. Он также поддерживает необходимую внутриклеточную концентрацию ионов Cl-, а в специализированных эпителиальных клетках обеспечивает их накопление до уровня, превышающего состояние электрохимического равновесия.
Например, в почке котранспортер Na+/K+/Cl- находится на апикальной мембране специализированных эпителиальных клеток, выстилающих просвет канальца толстой восходящей части петли Генле. Как иллюстрирует рисунок ниже, котранспортер играет критическую роль в реабсорбции NaCl из почечного фильтрата.
Мутации в гене, кодирующем изоформу котранспортера Na+/K+/Cl-, приводят к развитию аутосомного рецессивного заболевания, которое называется синдром Бартера. У больных наблюдается увеличение потерь воды и солей с мочой, снижение концентрационной функции почек и усиленное выведение из организма ионов Са2+. Такой эффект мутации в одной изоформе транспортера свидетельствует о том, что транспорт ионов через клетки осуществляется с одновременным участием многих типов транспортных белков и что нарушение функционирования одного из них отражается на функционировании остальных.
Действительно, мутации в других транспортных белках, которые функционируют совместно с котранспортером Na+/K+/Cl- и регулируют поглощение солей в почках, также приводят к развитию синдрома Бартера. К числу таких мутаций относятся мутации в К+- и в Cl--канале (CLC-K). Последний представляет собой часть базолатеральной системы проводимости Cl--ионов.
Еще одним примером Na+ зависимого вторичного транспортного белка является Na+/Mg2+-обменник. Поскольку ионы Mg2+ в больших концентрациях влияют на многие клеточные функции, внутриклеточная концентрация этого катиона тщательно контролируется буферным механизмом. Например, ионы Mg2+ конкурируют с ионами Са2+ за общие Ca2+/Mg2+-сайты активации/инактивации, расположенные на различных белках. Поскольку Mg2+ с небольшой скоростью проникает через плазматическую мембрану, он должен постоянно выводиться из цитоплазмы с тем, чтобы его концентрация поддерживалась в физиологических пределах. Выход Mg+ из клетки происходит, главным образом, с участием Na+/Mg+-обменника.
Предполагается, что этот обменник в физиологических условиях электронейтрален, и на каждый ион Mg2+, удаляемый из клетки, в нее закачиваются два иона Na+. Na+/Mg2+-обменник также является главной мишенью действия широко используемых антидепрессантов.
В зависимости от мембранного потенциала,
Na+/Са2+-обменник (NCX) функционирует в прямом и обратном направлениях.