• Для стабилизации концов хромосомы необходимы теломеры
• Теломера состоит из простых повторов, Ц+А-обогащенная цепь которых характеризуется последовательностью
Еще одной характерной чертой строения всех хромосом является теломера, которая «замыкает» их концы. Мы знаем, что теломера должна обладать особой структурой, поскольку концы хромосомы, образовавшиеся в результате разрыва, являются «липкими» и могут воссоединяться с другими хромосомами, в то время как естественные концы хромосом отличаются устойчивостью
При идентификации теломерной последовательности необходимо принимать во внимание два критерия:
• Последовательность должна быть расположена на конце хромосомы.
• Она должна придавать линейной молекуле ДНК устойчивость.
Методический подход, который позволил отбирать последовательности ДНК по функциональному критерию, был разработан опять-таки с использованием дрожжей. Все плазмиды, которые способны существовать в клетках дрожжей (и обладающие ARS- и CEN-элементами), представляют молекулы циркулярной ДНК. Линейные плазмиды нестабильны и деградируют.
Теломеры можно отбирать как последовательности, способные обеспечивать стабильность плазмид. Так можно идентифицировать фрагменты дрожжевой ДНК, локализованные на концах хромосом. Аналогичным образом, стабильность линейных форм дрожжевых плазмид обеспечивается концевыми участками природных линейных молекул ДНК — экстрахромосомной рДНК Tetrahymena.
Типичная теломера характеризуется простой повторяющейся структурой,
ГТ-богатая цепь которой выходит за пределы ЦА-богатой цепи.
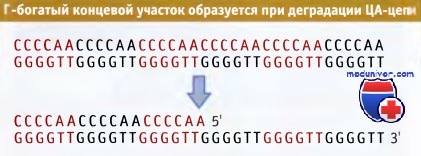
Г-богатый концевой участок образуется при ограниченной деградации ЦА-богатой цепи.
Теломерные последовательности были обнаружены в хромосомах самых разных представителей высших и низших эукариот. Для хромосом клеток растений и человека обнаружен одинаковый тип последовательности. Это свидетельствует о том, что в основе структурной организации теломеры лежит общий принцип. Каждая теломера состоит из протяженного участка коротких последовательностей, расположенных тандемно. В зависимости от организма он может содержать 100-1000 повторов.
Все теломерные последовательности могут быть представлены в виде общей формулы Сn(А/Т)m, где n > 1 и m = 1 - 4. На рисунке ниже представлен типичный пример.
Необычным свойством теломерной последовательности является продолжение ГТ-цепи в виде одиночной, длиной 14—16 оснований. По-видимому, Г-конец образуется за счет специфической ограниченной деградации ЦА-богатой цепи.
Теломерный участок реплицируется по специфическому механизму. Фермент теломераза представляет собой рибонуклеопротеид, который содержит матричную РНК, с той же последовательностью, что и в ЦА-богатой цепи. РНК присоединяется к теломере, образуя праймер, с которого начинается рост цепи с участием обратной транскриптазы, входящей в теломеразный комплекс. Процессивность фермента и число добавляемых повторов контролируются другими белками комплекса.
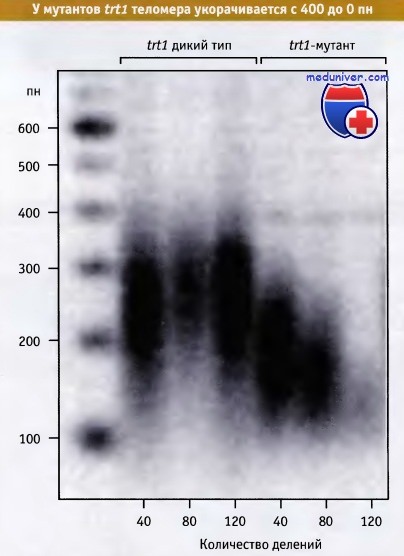
Поскольку репликация ДНК не может начаться с самого конца линейной молекулы, при репликации хромосомы количество повторов в теломере снижается. Это можно продемонстрировать, заблокировав активность теломеразы. Как следует из рисунка ниже, мутация по гену теломеразы приводит к укорачиванию теломерных участков с каждым делением клетки. Рисунок ниже показывает, что длина теломеры на протяжении 120 клеточных поколений снижается с 400 пн до нуля.
Способность теломеразы добавлять повторы к теломере при синтезе de novo, компенсирует их потерю за счет невозможности протекания репликации до конца хромосомы. Удлинение и укорачивание теломер находятся в динамическом равновесии. Если эти участки постоянно удлиняются (и укорачиваются), то их первичная последовательность не играет роли. Необходимо всего лишь, чтобы конец последовательности признавался подходящим субстратом для удлинения теломеры.
Теломеразная активность присутствует во всех делящихся клетках, и фермент обычно не обнаруживается в терминально дифференцированных, т. е. неделящихся клетках.
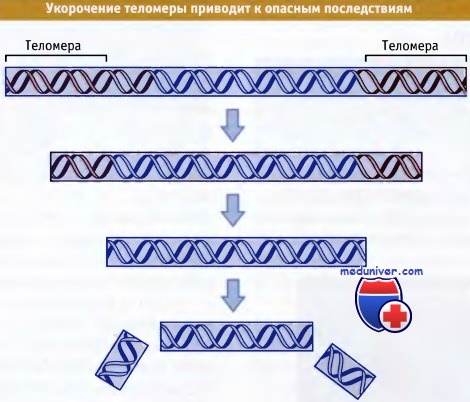
Мутации в теломеразе вызывают укорочение теломер после каждого цикла деления.
Полная потеря теломеры вызывает разрывы и перегруппировки хромосом.
В диком штамме дрожжей длина теломеры составляет около 350 пн,
однако у мутанта по гену trt1, кодирующего компонент РНК теломеразы, длина ее быстро сокращается до нуля.