Сопряжение трансляции и транслокации белков из ЭПР
• Взаимодействие между транслоконом и сигнальной последовательностью приводит к открытию канала и к началу транслокации.
• Детали механизма транслокации варьируют для разных белков.
После адресования и причаливания растущей цепи белка, связанной с рибосомой, должен начаться транспорт полипептида через мембрану. Большинство данных об этом процессе получено при исследовании сравнительно небольшого количества модельных белков в бесклеточной системе транслокации. Эффективность переноса зависит от скоординированности взаимодействий между белковой цепью, каналом и рибосомой. Эти взаимодействия меняют структуру канала и его связывание с рибосомой и образующейся цепью.
После высвобождения сигнальной последовательности из SRP на мембране эндоплазматического ретикулума (ЭПР), единственной силой, удерживающей рибосому на мембране, является ее взаимодействие с каналом. Такого взаимодействия оказывается недостаточно, чтобы начать перенос новообразующейся цепи, минуя этап ее узнавания на мембране ЭПР. Этот этап необходим, поскольку независимо от субстрата трансляции сами по себе рибосомы обладают не очень высоким, но существенным сродством к каналу.
Если бы для начала транслокации достаточно было только взаимодействия рибосомы с транслоконом, то также могли бы начать переноситься белки цитозоля. Такая возможность предотвращается следующим за причаливанием этапом, на котором канал должен узнать сигнальную последовательность.
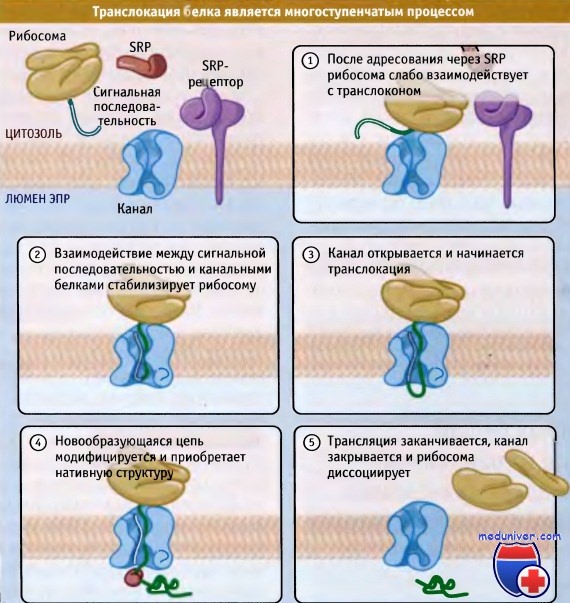
Рисунок ниже иллюстрирует изменения взаимодействия рибосомы с транслоконом при узнавании сигнальной последовательности. После причаливания и высвобождения SRP рибосома остается связанной с транслоконом лишь слабой связью. При этом сигнальная последовательность и остаток новообразующейся цепи ориентируются по направлению к цитозолю, но находятся в непосредственной близости от конца канала.
Вскоре после возобновления элонгации белковой цепи сигнальная последовательность узнается Sec61a. Предполагается, что для узнавания необходимо, чтобы сигнальная последовательность внедрилась в канал свернутой в виде петли, с N-терминальным концом, ориентированным в сторону цитозоля, а С-терминальным в сторону просвета ЭПР. При такой ориентации высвобождение белка с рибосомы в люмен ЭПР происходит через канал.
Узнавание сигнальной последовательности и начало ее поступления в канал представляют собой начальные этапы транслокации цепи. К ним, вероятно, также относятся смещение заглушки, закрывающей отверстие канала, и вставка в пору участка зрелой цепи (т. е. расположенного непосредственно после сигнальной последовательности). Как только это происходит, связь между рибосомой и каналом становится более прочной. После адресования все эти процессы, начиная с проникновения в пору сигнальной последовательности и до открытия канала, проходят очень быстро. Лишь небольшая дополнительная элонгация полипептида требуется, и описанные события завершаются задолго до того, как успеет синтезироваться большая часть белка.
Для того чтобы цепь секреторного белка могла начать транслоцироваться, необходимо, чтобы ее длина, расположенная вне рибосомы, составляла лишь около сорока аминокислот.
Если канал открылся, то большая часть секреторных белков переносится в люмен до окончания трансляции. Ранее было широко распространено мнение о том, что сила взаимодействия между рибосомой и транслоконом настолько велика, что эти компоненты прочно сцеплены друг с другом, чем предотвращается выход переносимой цепи в цитозоль. Однако многочисленными экспериментами было показано, что два компонента разделены промежутком, и, по крайней мере, в некоторых случаях часть цепи способна проходить через него в цитозоль.
Поскольку переносимые цепи имеют доступ в цитозоль, неизвестно, что побуждает их к переносу в люмен ЭПР, причем они действительно переносятся, а не просто падают. Одна возможность состоит в том, что взаимодействие с компонентами люмена ЭПР удерживает появившуюся цепь, не давая ей возможность выйти оттуда. Какой бы ни был этот механизм, считают, что субстрат транслокации предпочтительно мигрирует в люмен. Когда рибосома достигает стоп-кодона, цепь выходит в люмен, и канал закрывается, хотя точная последовательность этих событий пока неизвестна. Особенно непонятно, происходит закрытие канала после полного прохождения цепи или в результате диссоциации рибосомы.
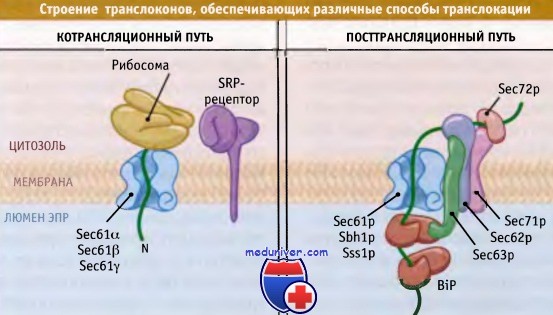
Хотя перечисленные процессы представляют собой основные этапы транслокации, они варьируют от субстрата к субстрату. Например, различные сигнальные последовательности по-разному взаимодействуют с белками каналов и открывают их по разным механизмам. В свою очередь, способ узнавания последовательности влияет на последующий биогенез некоторых белков. Для переноса ряда белков, наряду с Sec61 и TRAM, необходимы факторы ER. У определенных белков транслокация начинается с одних и тех же основных этапов, но позже проходит совсем по-другому. Например, некоторые белки на какое-то время выходят из зазора между рибосомой и транслоконом, при этом большие участки цепи находятся в цитозоле до момента обратного входа в канал.
Поэтому, хотя процесс транслокации обладает рядом общих существенных черт, было бы чересчур упрощенным думать, что она является единым механизмом переноса для разных белков через мембрану. Более вероятно, что специфический путь, по которому белок переносится через транслокон, оказывает впоследствии определенное влияние на его биологическую активность; однако меру этого влияния мы пока не можем оценить.
Элонгация новообразующейся цепи в люмен ЭПР не происходит до тех пор,
пока канал не узнал сигнальную последовательность и плотно не связал рибосому.
Вскоре после начала транслокации происходит модификация белковой молекулы,
и она приступает к приобретению нативной структуры.
Различные белки взаимодействуют с одним и тем же каналом,
обеспечивая котрансляционную или посттрансляционную транслокацию.