Механизм работы системы кальциевых сигналов клетки
• Почти во всех клетках ионы Са2+ служат вторичным мессенджером и выполняют регуляторные функции
• Са2+ непосредственно действует на многие белки-мишени, а также контролирует активность регуляторного белка калмодулина
• Концентрация Са2+ в цитозоле контролируется его депонированием в органеллы и высвобождением из них
Во всех клетках ионы Са2+ служат в качестве вторичного мессенджера, и, таким образом, они представляют собой более распространенную систему, чем цАМФ. Многие белки связывают кальций, что приводит к изменениям их ферментативной активности, субклеточной локализации или взаимодействия с другими белками и липидами.
К числу белков-мишенией, непосредственно регулируемых ионами кальция, относятся почти все классы сигнальных белков, описанные в настоящей главе, многочисленные ферменты, ионные каналы, насосы и сократительные белки. Среди них наибольшего внимания заслуживают актомиозиновые мышечные волокна, которые начинают сокращаться в ответ на изменение содержания в цитозоле кальция.
Хотя в большинстве внеклеточных жидкостей концентрация кальция составляет около 1 мМ, его внутриклеточная концентрация поддерживается на уровне 100 наномолей за счет совместного действия насосов и переносчиков, которые или выводят свободный кальций, или концентрируют его в эндоплазматическом ретикулуме или в митохондриях.
Передача сигнала с участием Са2+ происходит в тот момент, когда каналы эндоплазматического ретикулума или плазматической мембраны, селективные по кальцию, открываются, давая возможность катиону поступать в цитоплазму. К числу наиболее значимых каналов относятся потенциал-зависимые каналы плазматической мембраны клеток млекопитающих; Са2+-канал эндоплазматического ретикулума, который регулируется вторичным мессенджером, инозитол-1,4,5-трифосфатом (см. ниже); и потенциал-зависимый канал эндоплазматического (саркоплазматического) ретикулума мышц. Этот канал открывается в ответ на деполяризацию расположенной рядом плазматической мембраны, т. е. в результате процесса, известного как сопряжение возбуждения и сокращения.
Наряду с белками, активность которых регулируется при непосредственном связывании кальция, многие другие белки реагируют на ионы кальция, связываясь с калмодулином, широко распространенным, небольшим, 17 кДа, белком, чувствительным к кальцию. Для того, чтобы полностью активироваться, калмодулину необходимо связать четыре молекулы Са2+, причем связывание носит выраженный кооперативный характер, и профиль активации белка-мишени при этом имеет сигмоидную форму.
Обычно калмодулин связывается с мишенями в присутствии ионов кальция, но свободный, не содержащий кальция, калмодулин в некоторых случаях может оставаться в связанной но неактивной форме. Например, калмодулин является конститутивной субъединицей киназы фосфорилазы, которая активируется при связывании Са2+. У высших растений регуляция калмодулином носит иной характер. Калмодулин не экспрессируется в качестве самостоятельного белка, но вместо этого существует в качестве домена Са2+ зависимых белков. Еще одним исключением является аденилатциклаза, секретируемая патогенными бактериями Bordetella pertussis, которая вне клеток неактивна, но в клетках млекопитающих активируется свободным калмодулином.
При этом быстро образуется цАМФ, оказывающий сильное токсическое действие.
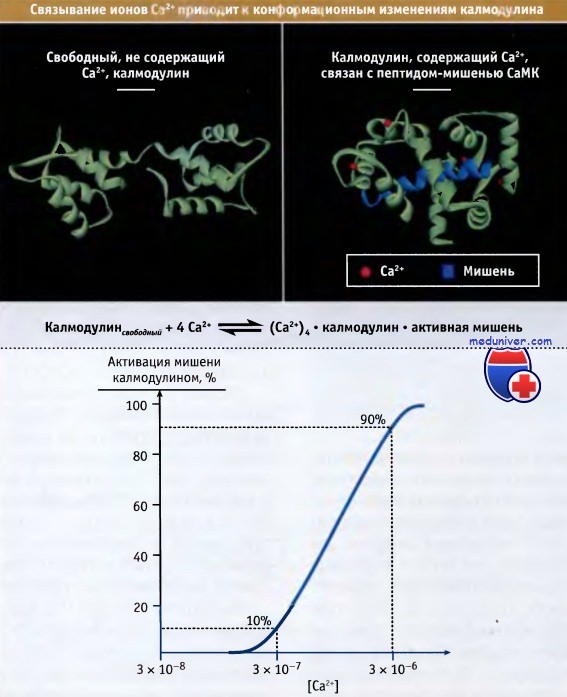
Диаграммы, иллюстрирующие кристаллическую структуру свободного (не содержащего кальция) калмодулина и белка, связанного с четырьмя ионами Са2+.
Заметны сильные конформационные изменения, которые претерпевает калмодулин после связывания Са2+. При связывания кальция с калмодулином, активность белков-мишеней меняется.
На нижнем рисунке показана активация мишени калмодулином как функция внутриклеточной концентрации свободных ионов Са2+.
Необходимость связывания четырех ионов Са2+ для изменения конформации белка приводит к кооперативной активации мишеней.
При увеличении концентрации кальция всего в 10 раз активность белка увеличивается с 10 до 90%.
Модели структуры построены по данным Protein Data Bank files 1CFD и 1MXE.