Синтез и строение белка: трансляция, посттрансляционные изменения
Белки, образуемые из полипептидов, — основные структурные компоненты тела человека, они катализируют большинство процессов метаболизма. Полипептидом называют цепь, состоящую из аминокислот, последовательность которых зависит от последовательности оснований в иРНК в соответствии с «генетическим кодом».
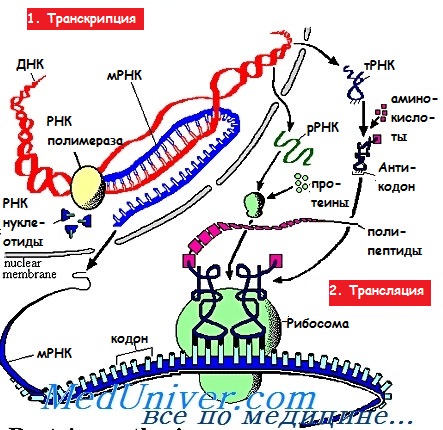
Каждая аминокислота представлена в иРНК одной или несколькими группа ми, называемыми триплетами кодонов, а их интерпретация в качестве полипептида носит имя трансляции. Трансляция иРНК происходит в рибосомах цитоплазмы в направлении от 5'- к З'-концу. Образуемые в результате полипептиды затем превращаются в белки.
Функциональные свойства белков обусловлены в основном их активными центрами, формирующимися при образовании третичной и четвертичной структур.
Генетический код для синтеза белка
Для трансляции необходимы молекулы тРНК, которые кодируют аминокислоты, соответствующие последовательности антикодона. Большинство аминокислот кодированы несколькими кодонами, исключение составляют триптофан и метионин (кодированы одним кодоном). Три из 64 возможных триплетных комбинаций А, Ц, Г и У в иРНК кодируют СТОП-сигналы: УГА, УАГ и УАА.
АУГ кодирует метионин и служит СТАРТ-сигналом, а также одновременно определяет амино-(N-)терминальный конец полипептида и формирует одну из трёх возможных рамок считывания. Генетический код митохондриальной ДНК немного отличается.
Трансляция при синтезе белка
Инициация. Малая субъединица рибосомы, содержащая несколько факторов инициации и кодирующая метионин метионил-тРНК, связывается с 5'-кэпом иРНК, а затем начинает скольжение вдоль молекулы иРНК и, достигнув первой АУГ-последовательности, сцепляется с ней. При этом происходит высвобождение факторов инициации, а большая субъединица рибосомы прикрепляется к малой, после чего запускается процесс трансляции.
Большая субъединица рибосомы имеет в своём составе два специализированных участка, известных как А-сайт (аминоацил-тРНК-связывающий участок) и Р-сайт (пептидил-тРНК-связывающий участок). В конце инициации к Р-сайту прикреплена заряженная мет-тРНК, антикодон которой сцеплен с АУГ-кодоном, в то время как А-сайт свободен.
Элонгация. В соответствии с расположенным рядом кодоном иРНК к А-сайту при помощи растворимого фактора элонгации 1 (EF1) прикрепляется аминоацил-тРНК. Под воздействием пептидилтрансферазы возникает пептидная связь между аминогруппой (-NH2) аминокислоты на А-сайте и карбоксильной группой (-СООН) на Р-сайте, при этом первая тРНК отделяется.
Фермент транслоказа обеспечивает отделение свободной тРНК, движение рибосомы вдоль молекулы иРНК, а также перенос растущей пептидной цепочки от А-сайта к Р-сайту. Для осуществления данного процесса необходим фактор EF2.
Митохондриальные иРНК транслируются при помощи митохондриоспецифических тРНК.
Терминация. Элонгация продолжается до тех пор, пока на пути рибосомы не встретится один из терминирующих (СТОП) кодонов, которые распознают благодаря мультивалентным факторам освобождения (терминации, RF). При этом специфичность пептидилтрансферазы изменяется таким образом, что к | белку прикрепляется молекула воды. Затем рибосома отделяется и распадается на составляющие её субъединицы, освобождая готовый полипептид.
Для синтеза полипептида, длина которого составляет примерно 400 аминокислот, в среднем необходимо до 20 с.
Как только одна рибосома освобождает кэп иРНК, к нему присоединяется следующая, формируя полирибосому или полисому. Время жизни иРНК составляет несколько часов.
Строение белка
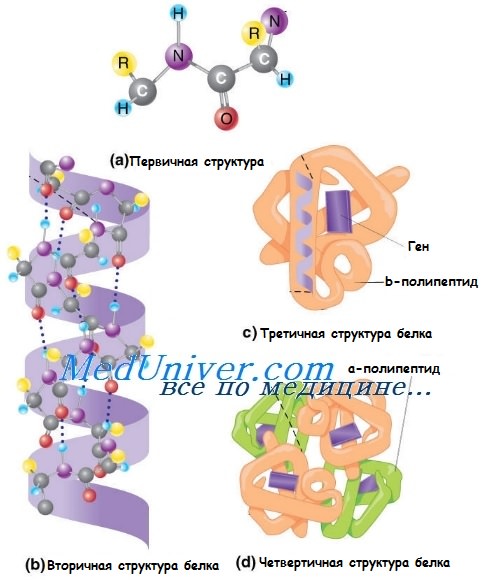
Аминокислотная последовательность полипептида обусловливает его первичную структуру.
Вторичная структура белка представляет собой трёхмерное расположение частей полипептида: а-спираль, коллагеновая про-а-спираль или бета-складочная конформация.
Третичная структура белка образуется при самоукладке всех вторичных структур полипептида.
Четвертичная структура белка — окончательная конформация мультимерного белка, такого, как, например, гемоглобин, который состоит из двух а-глобулиновых и двух бета-глобулиновых мономеров, а также молекулы гема и атома двухвалентного железа. Коллагеновые волокна состоят из трёхспиральных нитей, которые образованы тремя про-а-спиралями.
Чаще всего структуру белка поддерживают дисульфидные мосты, которые возникают между серосодержащими остатками цистеина расположенных рядом цепей, в то время как ферментативные свойства зависят прежде всего от распределения заряженных групп.
Посттрансляционные изменения белка
К посттрансляционным изменениям относят удаление N-терминального метионина и дробление. Объединение происходит между одинаковыми и разными полипептидами, а также между простетическими группами, к которым относят гем.
Полипептиды, предназначенные для внеклеточной (экстрацеллюлярной) секреции, вначале подвергаются гликозилированию в эндоплазматическом ретикулуме и аппарате Гольджи. Для их отбора имеет значение сигнальный пептид, расположенный около N-конца, который связывается с сигналраспознающей частицей, состоящей из цитоплазматической 7SL-PHK и шести специфических белков.
При помощи сигнального пептида происходит связывание с мембранным рецептором эндоплазматического ретикулума. Сразу после синтеза полипептид переносится через мембрану; как только появляется его С-конец (карбоксильная часть), сигнальный пептид отделяется. Затем полипептиды в составе везикул, которые отпочковываются от эндоплазматического ретикулума, попадают к аппарату Гольджи.
Гликозилированию обычно подвержен N-конец пептида. При этом происходит присоединение олигосахарида к NH2-rpynne боковой цепи аспарагина (например, при производстве антител и лизоцима). Иногда олигосахарид присоединяется к ОН-группе боковой цепи серина, треонина или гидроксилизина (например, секретируемые антигены групп крови системы АВО).
Среди других модификаций различают гидроксилирование лизина и пролина, играющее важную роль в образовании про-а-спиралей коллагена, сульфатацию тирозина, служащую сигналом к компартментализации (пространственному разделению), а также липидизацию остатков цистеина и глицина, необходимую для их прикрепления к фосфолипидной мембране.
Ацетилирование лизина в гистоне Н4 влияет на его связывание с ДНК. Протеинкиназы фосфорилируют остатки серина и тирозина и могут регулировать ферментативную способность энзимов, как в случае с каскадной сигнальной трансдукцией протоонкогенов.
Медицинское значение понимания синтеза белка
В основе механизма возникновения болезни клеточных включений лежит недостаточность гликозилирования лизоцима. Рицин, входящий в состав бобов клещевины обыкновенной (Ricinus communis), блокирует EF2, а дифтерийный токсин — транслоказу.
Мишенью для большого количества антибиотиков служат процессы трансляции, характерные для прокариот. Например, эритромицин разрушает транслоказу, хлорамфеникол взаимодействует с пептидилтрансферазой, тетрациклин препятствует связыванию аминоацил-тРНК, пуромицин по своей структуре сходен с аминоацил-тРНК, а стрептомицин связывается с малой субъединицей бактериальных рибосом. При этом митохондрии человека имеют эволюционное родство с бактериями, поэтому некоторые антибиотики оказывают влияние и на митохондриальные функции.