• Основные клеточные фосфолипиды синтезируются в мембране ЭПР, главным образом, со стороны цитозоля.
• Регуляция синтеза новых липидов контролируется локализацией ферментов, участвующих в этом процессе.
• Биосинтез холестерина регулируется протеолизом фактора транскрипции, который интегрирован в мембрану ЭПР.
Наряду с транслокацией и подготовкой белков к секреторному процессу, эндоплазматический ретикулум (ЭПР) функционирует как место первичного образования клеточных фосфолипидов. Эти молекулы синтезируются в мембране эндоплазматического ретикулума (ЭПР) и затем распределяются по отдельным мембранам и органеллам внутри клетки. К их числу относится плазматическая мембрана, митохондрии и органеллы секреторного пути.
При возникновении необходимости клетка должна обладать способностью увеличивать количество мембран. Наиболее очевидно это проявляется при удвоении плазматической мембраны и всех органелл, которое происходит в ходе клеточного цикла. Наряду с этим, при гиперфункции, отдельные органеллы могут увеличиваться в размерах. Например, при созревании предшественников В-лимфоцитов в плазматические клетки, секретирующие антитела, ЭПР сильно увеличивается, что соответствует потребностям усиленного транспорта белков по секреторному пути.
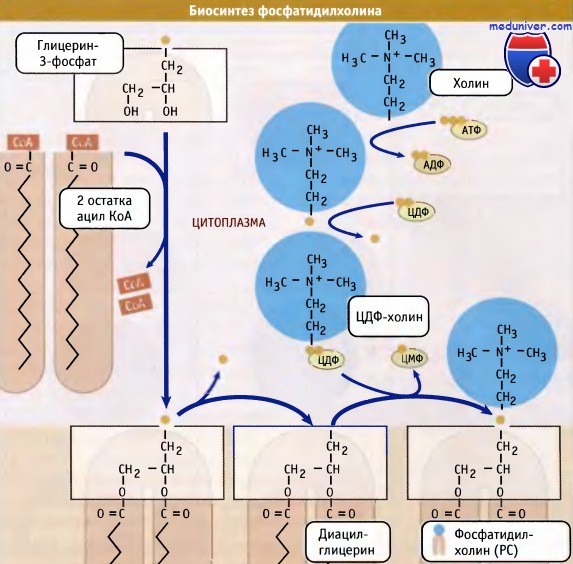
Образование фосфолипидов из растворимых предшественников de novo происходит, главным образом, на мембране ЭПР со стороны цитоплазмы, в ходе процесса, называемого цикл Кеннеди. Как показано на при росте мембраны ЭПР две молекулы жирных кислот, связанные с ацетил КоА, реагируют с глицерин-3-фосфатом, образуя диацилглицерин (ДАТ). В отличие от своих предшественников, ДАГ достаточно гидрофобен для того, чтобы интегрироваться в мембрану ЭПР.
Попав в мембрану, ДАГ взаимодействует с «головой» будущей молекулы фосфолипида. Как показано на рисунке ниже, основные фосфолипиды различаются, главным образом, природой «головы» молекулы. Все они образуются при присоединении «головы» к ДАГ. Вначале происходит ее фосфорилирование, а затем присоединение к цитидиндифосфату (ЦДФ). После этого «голова» с одной из фосфатных групп переносится на ДАГ, и синтез завершается.
Присоединение головной группы холина к ЦЦФ с образованием фосфатидилхолина иллюстрирует использование клеткой субклеточной локализации белков для регуляции процессов биосинтеза. В этой реакции участвует фермент цитидилтрансфераза (ЦТ), и она контролирует скорость всего процесса синтеза фосфатидилхолина. В клетке ЦТ находится в двух различных формах: в неактивной, в цитозоле, и в активной, с цитозольной стороны мембраны ЭПР.
Механизм, который обеспечивает существование двух этих форм фермента, представляется не вполне ясным. Очевидно, однако, что условия внутри клетки, которые сигнализируют о необходимости усиления синтеза фосфатидилхолина, могут контролировать переход ЦТ из одной формы в другую. Например, увеличение в клетке концентрации свободного ДАТ или жирных кислот вызывает миграцию ЦТ на мембрану ЭПР, где эти компоненты реагируют с холином. В то же время, когда в клетке содержится много фосфатидилхолина, ЦТ локализуется в цитозоле. Таким образом, предотвращается синтез фосфатидилхолина.
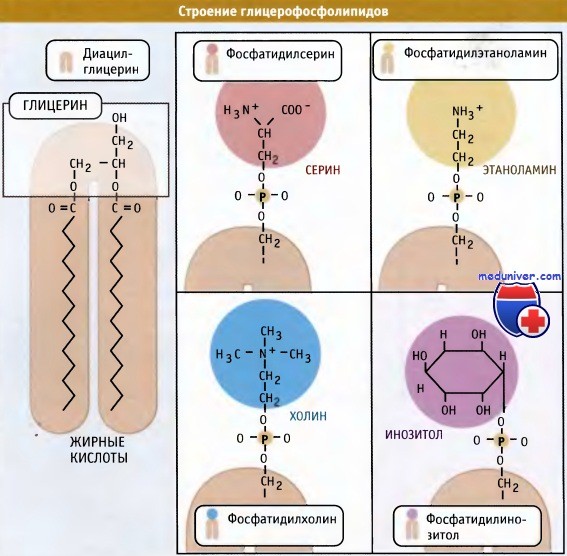
Наряду с фосфатидилхолином, который является наиболее распространенным мембранным фосфолипидом, в цикле Кеннеди также образуются фосфатидилэтаноламин и фосфатидилинозитол. У дрожжей S. cerevisiae синтезируется фосфатидилсерин. У животных, однако, этот фосфолипид образуется за счет другой реакции, которая не требует участия ЦЦФ, однако также протекает с цитозольной стороны мембраны ЭПР.
Интересная особенность синтеза фосфатидилэтаноламина заключается в том, что хотя этот фосфолипид образуется в цикле Кеннеди, он также может синтезироваться в митохондриях. В этих органеллах он образуется при модификации фосфатидилсерина, который образовался в ЭПР. Образование фосфатидилэтаноламина в клеточных компартментах, отличающихся от тех, в которых синтезируется его предшественник, предполагает существование механизма транспорта липидов между двумя компартментами.
Имеются данные, позволяющие предполагать, что этот транспорт осуществляется посредством специального субдомена ЭПР, который называется мембраной, связанной с митохондрией (МСМ). МСМ представляет собой участок ЭПР, который физически ассоциирован с митохондрией. Он характеризуется плоской поверхностью и тесным контактом с органеллой. В МСМ находятся ферменты синтеза фосфатидилсерина. Возможно, что область контакта мембраны с митохондрией обеспечивает быстрый перенос фосфатидилсерина в органеллу, хотя механизм этого переноса остается неясным.
Наряду с участием в синтезе фосфолипидов, эндоплазматический ретикулум (ЭПР) является местом синтеза стеринов. К числу основных стеринов мембран клетки относится холестерин; его биосинтез осуществляется в несколько этапов. Первые несколько реакций происходят в цитозоле, а остальные в мембране ЭПР. Также в ЭПР находятся элементы, необходимые для регуляции образования холестерина. Ключевыми медиаторами этого регуляторного пути являются белки, связывающиеся с регуляторным стеринреспонсивным элементом (SREBP).
Эти белки обычно интегрированы в мембрану эндоплазматического ретикулума (ЭПР) и со стороны люмена содержат небольшую петлю, соединяющую два трансмембранных домена, N- и С-концевые участки которых ориентированы в сторону цитозоля. Также в ЭПР находится белок SCAP (белок, активирующий разрезание SREBP). Этот белок контролирует уровень внутриклеточного холестерина. Когда возникает необходимость в повышенном синтезе холестерина, SCAP транспортирует SREBP из ЭПР в аппарат Гольджи. Там происходит разрезание SREBP, и N-терминальный участок молекулы высвобождается в цитозоль. (В реакции участвуют те же протеазы, которые расщепляют АТФ 6 при возникновении стрессорных условий в ЭПР. Затем N-терминальный домен SREBP мигрирует в ядро, где действует как фактор транскрипции, активируя экспрессию генов в каскаде реакций биосинтеза холестерина.
При объединении водорастворимых цитоплазматических компонентов (глицерин-3-фосфата, ацил КоА и полярной головки) образуется молекула фосфолипида.
Новая растворимая в липидах молекула включается в мембрану.
Основные мембранные фосфолипиды различаются по составу головок.