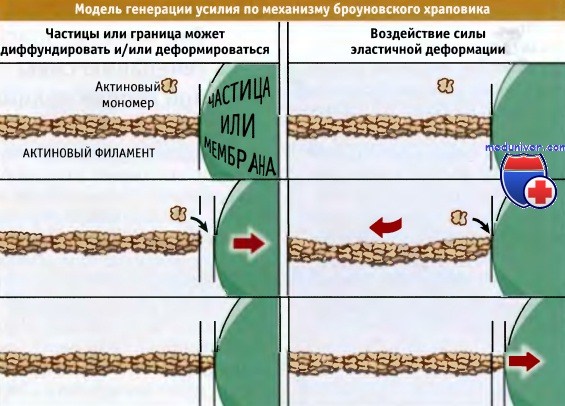
Каким образом добавление актиновых мономеров в филамент генерирует силу, необходимую для протрузии мембраны? Предложены две модели «броуновского колеса», описывающие, как в результате полимеризации актина генерируется усилие и происходит смещение частицы или границы мембраны.
Обе модели предполагают наличие некоего компонента (актинового филамента или транспортируемой частицы), который находится в беспорядочном температурном (броуновском) движении. Также они постулируют, что концентрация актиновых мономеров превышает критическую, так что может происходить элонгация филамента. Однако модели различаются в зависимости от природы диффундирующего компонента (актиновый филамент или частица).
Согласно одной модели, актиновый филамент зафиксирован, обладает достаточной жесткостью и не может изгибаться. Частица (например, везикула) или мембрана (например, перемещающийся участок мембраны подвижных клеток) благодаря броуновскому движению постоянно меняет свое положение. Когда оперенный конец филамента находится в контакте с мембраной, его элонгация не происходит, поскольку свободные мономеры актина не могут присоединиться к оперенному концу.
При движении частицы или мембраны за счет диффузии между ней и оперенным концом филамента образуется зазор. Когда этот зазор становится достаточно большим, происходит присоединение мономера и тем самым предотвращается диффузия частицы в обратном направлении. В результате предупреждения обратной диффузии и заполнения появляющихся промежутков между оперенным концом актинового филамента и частицей, изменяется характер движения частицы. Она начинается двигаться в определенном направлении.
Скорость движения частицы зависит от того, насколько быстро она способна диффундировать, и от величины вероятности добавления нового актинового мономера (что, в свою очередь, определяется концентрацией актина и размером области, доступной для элонгации). При этом чем выше скорость диффузии частицы, тем быстрее происходит элонгация оперенного конца, и тем быстрее частица продвигается вправо.
Вторая модель учитывает способность филаментов и частиц участвовать в броуновском движении. Поскольку интенсивность движения пропорциональна величине частицы (более крупные частицы диффундируют медленнее, чем более мелкие), частицы, по размерам превышающие актиновые филаменты, считаются практически неподвижными. Актиновые филаменты неспособны к быстрой диффузии, однако за счет температурного эффекта они могут сгибаться.
Таким образом, за счет этого может открываться пространство между частицей и филаментом, в результате чего становится возможным добавление мономера к оперенному концу филамента. Согнувшийся филамент находится в напряженном состоянии и стремится выпрямиться. Это сила эластичности действует на частицу, которая при достаточной величине силы по мере выпрямления филамента перемещается.
Генерация силы проиходит за счет реакции полимеризации и не требует энергии гидролиза АТФ. Гидролиз АТФ требуется для реутилизации актиновых субъединиц, с тем чтобы процесс мог произойти снова. Без реакции гидролиза и реутилизации мономера процесс происходит только один раз.
Две модели, описывающие, каким образом полимеризация актиновых филаментов, присоединенных к оперенному концу,
генерирует усилие, достаточное для продвижения мембраны или везикулы.