Сигнальный транспорт и неизбирательный поток белков
• Для растворимых секреторных белков, особенно секретирующихся в больших количествах, не всегда нужны специфические сигналы экзоцитоза
• Необходимость сигналов сортировки может ограничиваться мембранными белками и рецепторами, особенно теми, которые направляются к некоторым органеллам, например в лизосомы
• Некоторые растворимые белки несут сигналы, позволяющие взаимодействовать с рецепторами, которые участвуют в транспорте этих белков в лизосомы
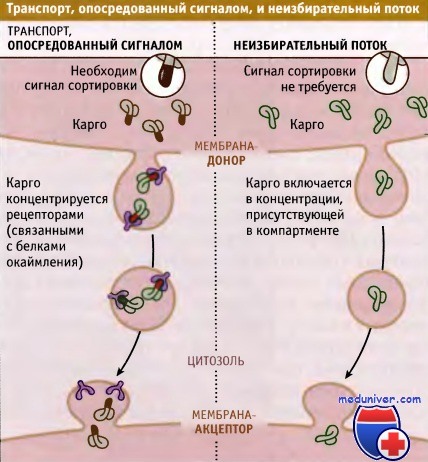
Существуют два представления о путях транспорта белков по механизму эндоцитоза и экзоцитоза. Это сигнальный транспорт и неизбирательный поток. При сигнальном транспорте каждый белок-карго содержит одну или несколько коротких последовательностей, определяющих его местоположение на пути транспортировки.
Наличие или отсутствие этих «последовательностей сортировки» (также называемых сигналами сортировки) определяет, остается ли белок в составе данной органеллы в качестве резидентного компонента, или транспортируется в везикуле в следующую органеллу, в которой он может остаться или следовать дальше по назначению. Если на каждом этапе требуется особый сигнал, то каждый белок должен содержать несколько последовательностей соответственно числу транспортных этапов, которые он должен преодолеть, чтобы дойти до пункта назначения (например, до плазматической мембраны при экзоцитозе или до лизосомы при эндоцитозе).
Для растворимых белков, находящихся в люмене везикулы и в компартментах, проблема транспорта стоит более остро, чем для мембранных белков. С точки зрения топологии растворимые белки находятся «вне клетки». Для того чтобы их транспорт полностью определялся сигналом, они должны связываться с мембранными рецепторами, которые, в свою очередь, связаны с компонентами окаймления, участвующими в селективном накоплении карго в везикулах.
Если каждому классу растворимых белков на определенном этапе транспорта необходим свой рецептор, то в каждый момент времени для транспорта белков потребовались бы десятки и сотни рецепторов.
Молекулы карго или концентрируются в везикулах с участием рецепторов,
которые связываются с сигналом сортировки на карго (транспорт, опосредованный сигналом),
или включаются в везикулы в той концентрации, в которой они находятся в компартменте или мембране (неизбирательный поток).
Второе представление является альтернативой первому и упрощает проблему. Модель неизбирательного потока определяет «пути по умолчанию» как не требующие существования у белков сигналов сортировки для транспорта между компартментами. Экзоцитоз и эндоцитоз должны рассматриваться как пути по умолчанию. Например, для поступления в везикулы при экзоцитозе секретируемые белки или белки плазматической мембраны не нуждаются в сигнале сортировки. Эти белки должны присутствовать в отпочковывающихся от ЭПР везикулах в той же концентрации, что и в люмене ЭПР. Механизм неизбирательного потока не включает сигнала сортировки, который используется для концентрации белка в формирующейся везикуле.
Сигнал требуется только в том случае, когда необходимо приостановить движение белка по пути экзоцитоза (например, при его удержании в аппарате Гольджи) или перевести его на другой путь (например, на путь транспорта в лизосомы).
Модель неизбирательного потока объясняет, каким образом бактериальный белок b-лактамаза, введенный в ооцит Xenopus, транспортируется из ЭПР на поверхность клетки. У бактерий отсутствуют внутренние мембраны, подобные тем, которые находятся в клетках эукариот. Поэтому бактериальные белки, по-видимому, не имеют сигнала экспорта из ЭПР. Неизбирательным потоком также объясняется высокая скорость экспорта белков из специализированных секреторных клеток, таких как ацинарные клетки поджелудочной железы.
Скорость секреции и многообразие секретируемых белков должны были бы превышать возможности клетки обеспечить рецептором каждый белок на каждом этапе транспорта.
Транспорт белков происходит по механизму как сигнального транспорта, так и неизбирательного потока. Механизм неизбирательного потока впервые был предложен для описания экзоцитоза. Для многих белков, выходящих из ЭПР к клеточной поверхности через аппарат Гольджи, сигналы, по-видимому, не нужны. Однако существуют данные, что некоторые мембранные и секреторные белки, выходящие из ЭПР, несут сигналы экспорта или используют дополнительные белки, которые допускают их селективный выход.
Данные, имеющиеся в отношении белков, которые транспортируются по пути эндоцитоза, позволяют предполагать, что для некоторых этапов этого процесса основным механизмом является сигнальный транспорт. Например, сигналы сортировки, расположенные на цитоплазматическом участке мембранных белков, обеспечивают их интернациализацию в плазматическую мембрану. Однако интернализованные растворимые макромолекулы транспортируются из эндосом в лизосомы, не связываясь с рецепторами.
Если концентрация молекул небольшая, то сигналы увеличивают эффективность интернализации. Небольшие молекулы, добавленные к среде, окружающей нефагоцитирующие клетки в культуре, интернализуются в жидкую фазу (т. е. не связанную с рецепторами) менее эффективно, чем при связывании с мембранными рецепторами. Связывание сигналов поглощения с рецепторами существенно увеличивает эффективность захвата белков (в 1000 раз) за счет концентрирования молекул в везикулах. Однако при высокой концентрации молекул значительные количества белков интернализуются даже без связывания с рецепторами.
Эти соображения также относятся к экспорту белков из эндоплазматического ретикулума (ЭПР). Неселективный поток растворимых белков тоже достигает значительной величины, когда транспортные везикулы обладают большим диаметром, например при макропиноцитозе, происходящем с участием плазматической мембраны.